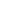|
| |
Cylindrospermopsis raciborskii (Woloszynska) Seenayya et Subba Raju, 1972
Цилиндроспермопсис Рациборского |
Систематическое положение (Kingdom, Phylum, Class, Order, Family, Species):
Bacteria » Cyanobacteria » Cyanophyceae » Nostocales » Nostocaceae » Cylindrospermopsis raciborskii
Цилиндроспермопсис Рациборского - Cylindrospermopsis raciborskii (Woloszynska) Seenayya et Subba Raju, 1972
Русское и английское названия. Цилиндроспермопсис Рациборского, Cylindrospermopsis raciborskii
Синонимы. Anabaena raciborskii Woloszynska, Anabaenopsis raciborskii (Wolosz.) Elenkin
Нативный ареал. Цианобактерия C. raciborskii известна как пантропический вид. Найдена в Индонезии, Индии, Таиланде, Японии, Филиппинах, Египте, Сенегале, Бразилии, Кубе, Чили, Уругвае, Австралии, Новой Зеландии, Флориде.
Современный ареал (мировой и конкретнее в России). Как инвазионный вид встречается в Венгрии, Австрии, Польше, Испании, Словакии, Германии, Франции, США, Канаде, Чехии. Отмечен также в Украине, Белоруссии, Молдавии, Западной Словакии, Греции, но наиболее широко распространен в тропической (включая субтропики) зоне Евразии, Австралии, Африки, Северной и Южной Америки. Вид отмечали в водоемах Узбекистана, Казахстана, Туркмении (до 50о с.ш.). Самые северные находки – водоемы Германии в пределах 53–54о с.ш. В России отмечен в Финском заливе в губах Копорская и Лужская. Известен еще на юге европейской части: в нижнем течении р. Дон, в Таганрогском заливе Азовского моря, в дельте Волги и Северном Каспии, в Веселовском и Цимлянском водохранилищах. Зарегистрирован в оз. Неро (бассейн Верхней Волги, 57,06о – 57,12ос.ш.), где отмечено вызванное им «цветение» воды. Найден в водоемах низовья р. Амударьи, в водоеме-охладителе Запорожской АЭС (Padisбk, 2003)
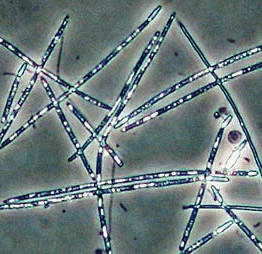
 Инвазионная часть ареала цилиндроспермопсис рациборского (Cylindrospermopsis raciborskii) в северной Евразии. 1- места находок вида (см. текст), 2 широкое распространение на начальном участке северного инвазионного коридора, 3- северный коридор инвазии;
4 - широкое распространение, 5- страны присутствия. Стрелками указано направление распространения в Европе
© Проект РНФ, № 16-14-10323 (Рук. В.Г. Петросян), ИПЭЭ РАН
Пути(коридоры) и векторы (способы) интродукции.
Экспансия в водоемы умеренной зоны пресноводной планктонной цианобактерии Cylindrospermopsis raciborskii, изначально описанной как Anabaena raciborskii для водоемов субтропической и тропической зон (Woloszynska, 1912), происходит чрезвычайно масштабно. Уже в 1997 г. Ю. Падишак (Padisak, 1997), анализируя обширный материал, отмечала высокую адаптивную способность вида к экспансии, его широкую экологическую валентность. Этому способствует целый ряд характеристик вида. Несмотря на предпочтение высоких температур, C. raciborskii толерантен и к низким (Briand et al., 2004; Bonilla et al., 2012). Эврифотен, устойчив к условиям высокой перемешиваемости водных масс (Padisak, 1997; Briand et al., 2004). Диазотроф: способен к азотфиксации, а также использованию растворенных форм азота, с предпочтением аммония (Saker, Neilan, 2001). Имеет высокое сродство к фосфору и может его запасать больше других Cyanoprokaryota (Istvanovics et al., 2000). Способен к вертикальной миграции за счет регулирования количества газовых вакуолей, устойчив к выеданию зоопланктоном (Padisak, 1997). Наиболее дискутируемые гипотезы продвижения в северные широты на фоне глобального потепления: селекция с формированием экоморф (Сhonudomkul et al., 2004) и широкая физиологическая толерантность вида в целом (Briand et al., 2004). Массовое образование акинет в подавляющем большинстве популяций умеренной зоны рассматривается как адаптационный механизм переживания зимнего периода (Padisak, 1997, 2003).
Местообитание. Обитает в толще воды пресноводных и слабосолоновато водных водоемов - озер, прудов, водохранилищ, на рисовых полях, реже в реках, эстуариях, опресненных морских заливах. До середины XX века указывался главным образом в тропических районах как обычный возбудитель "цветения" воды. В последние десятилетия отмечается проникновение вида в умеренный пояс, где он также образует "цветение" воды в теплые сезоны года (летом, осенью). Потенциальный азотфиксатор. Теплолюбивый. Ограничивающим фактором является температура воды ниже 16о С. Однако возможно образование клонов, адаптированных к холодной воде.
Особенности биологии. Cylindrospermopsis raciborskii — это нитчатая цианобактерия, имеющая способность фиксировать азот путем трансформации атмосферного азота (N2) в аммоний (NH3). При недостатке соединений азота в среде вдоль трихомы из вегетативных клеток и акинет появляются гетероцисты. Гетероцисты неспособны обычно к делению и росту. В них разрушается фотосистема II, соответственно не идет фотосинтез и не образуется внутренний кислород. От внешнего молекулярного кислорода гетероцисты защищены двумя толстыми дополнительными оболочками. Внутренняя состоит из гидроксилированных гликолипидов, наружная — из полисахаридов. Дифференцировка клетки трихома в сторону образования гетероцисты контролируется NtcA ДНК-связывающим белком, при этом повышается экспрессия гена hetR. Его действие модулируют продукты генов hetF и patA. Этот процесс может ингибироваться наличием аммония в среде. Для гетероцист отмечена высокая оксидазная активность, нейтрализующая прошедший кислород. Гетероцисты связаны с соседними клетками в трихоме с помощью плазмодесм, по которым идет транспорт связанного азота из гетероцисты, а органических соединений в неё. В гетероцисте молекулярный азот с помощью нитрогеназы переводится в аммоний, затем при участии глутамин-синтазы (ГС) аммоний с глутаматом превращаются в глутамин. Поскольку глутамат-синтаза (ГОГАТ) в основном представлена в вегетативных клетках, образованный глутамин передается туда из гетероцист, где он посредством ГОГАТ превращается в глутамат. Глутамат поступает из вегетативной клетки в гетероцисту и цикл замыкается. Из глутамата и глутамина образуются остальные аминокислоты в вегетативных клетках. Связанный азот запасается в цианофициновых гранулах. Cylindrospermopsis raciborskii фиксирует углерод в вегетативных клетках путем фотосинтеза, который переходит в форму углеводов (глюкоза/сахароза). При неблагоприятных условиях из вегетативных клеток образуются акинеты. Акинеты (от греч. Akinetos — неподвижный) — покоящиеся клетки цианобактерий с утолщённой оболочкой, большим количеством запасных питательных веществ и пигментов. Они служат для переживания неблагоприятных условий (устойчивы к пониженным температурам и высушиванию).
Влияние вида (на другие виды, экосистемы включая лесную и агроценозы, здоровье человека).
Эта цианобактерия опасна тем, что продуцирует чрезвычайно токсичные циклические пептиды (цилиндроспермопсин и его производные). Цилиндроспермопсин угнетает функцию печени, почек и других органов, ингибирует синтез белка и может приводить к летальному исходу. Для человека он опасен даже при контакте с кожей или употреблении отравленной пресноводной рыбы. Особенно опасен при попадании в резервуары с питьевой водой. Первое описание проблем со здоровьем, вызванное этой Cylindrospermopsis raciborskii, касается Австралии (Bourke et al. 1983). Болезнь проявлялась в виде гепатита. Цианобактерия содержит мало полиненасыщенных жирных кислот, что приводит к сокращению передачи вещества и энергии по трофическим цепям и к угнетению развития выше стоящих трофических уровней, снижает разнообразие водных экосистем. Вид относится к высоко вредным и токсичным инвазионным видам (Antuneset al., 2015).
|
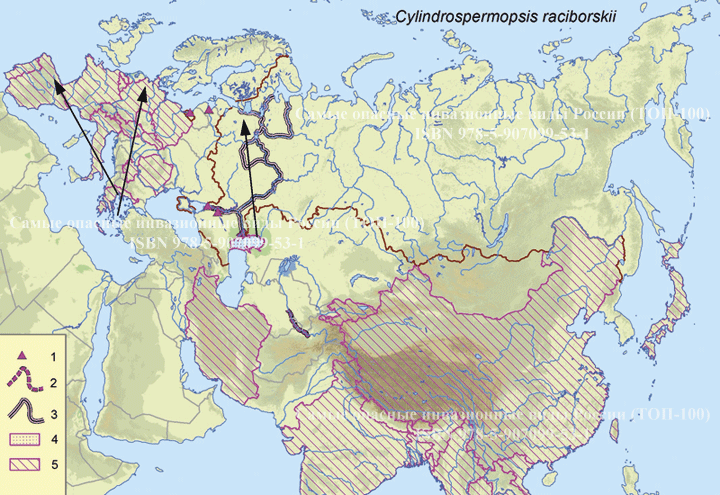
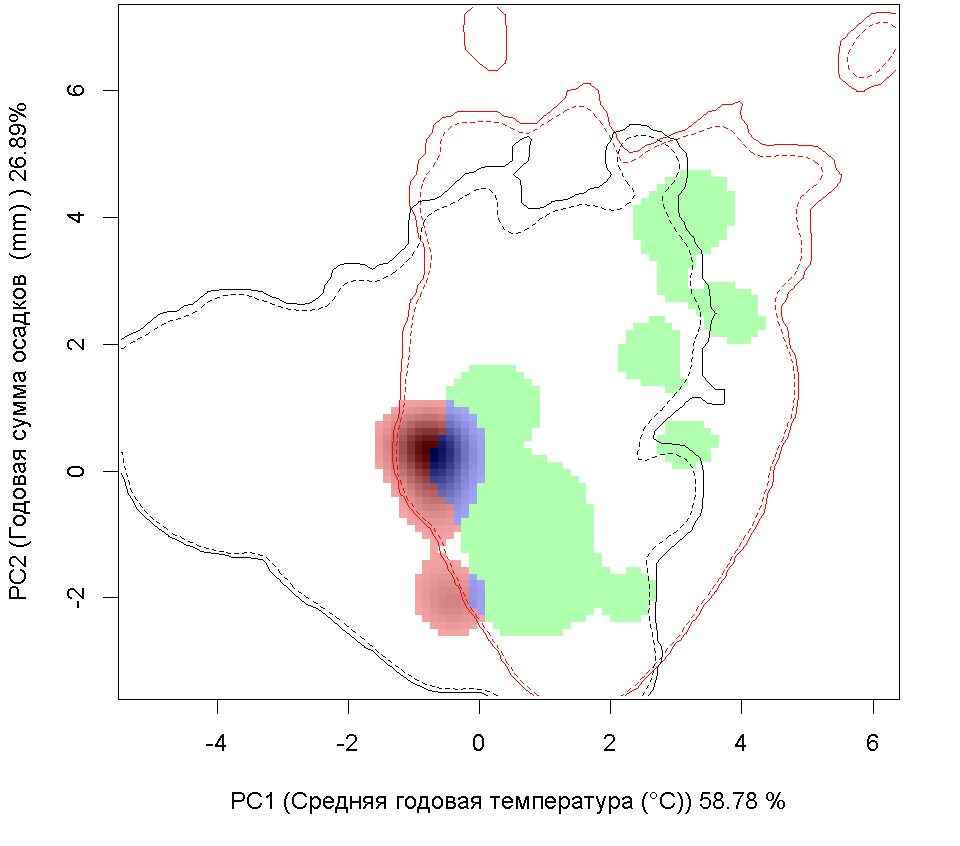
Графическое представление перекрывания ниш нативной и инвазионной частей ареалов, а также при сценариях изменения климата
|
|
Текущий климат
|
|
Нативная часть
|
Инвазионная часть
|
|
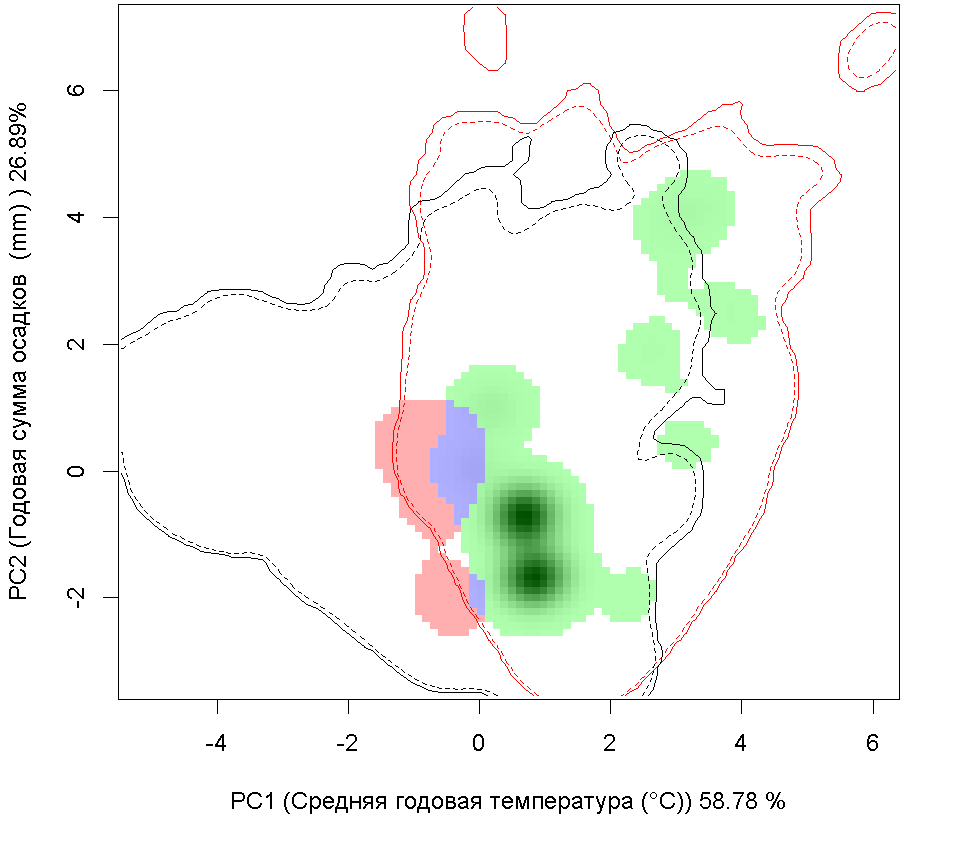 A A
|
 B B
|
| Графическое представление перекрывания ниш нативной (A) и инвазионной (B) частей ареалов вида, где сиреневый цвет – зона стабильности, розовый цвет – зона расширения, зеленый цвет – зона «неиспользования». Сплошные и пунктирные линии показывают 100% и 90% области доступной среды в нативной (черные линии) и инвазионной (красные линии) частях ареалов, которые использовались для анализа перекрывания ниш. |
|
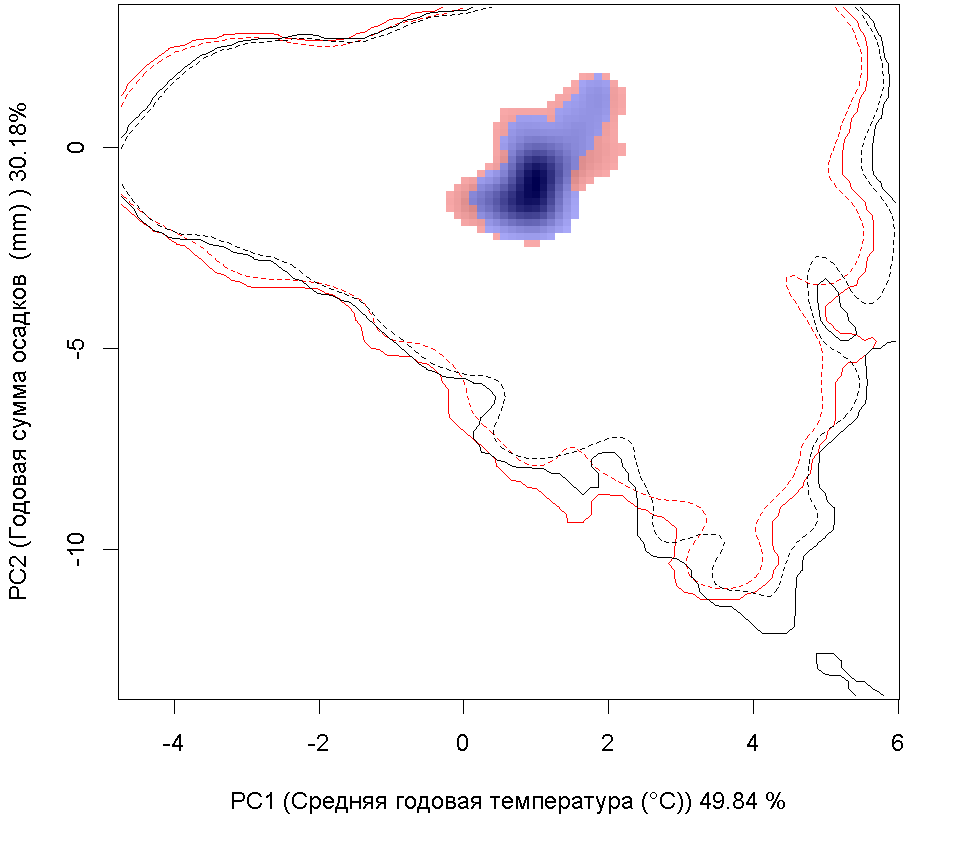
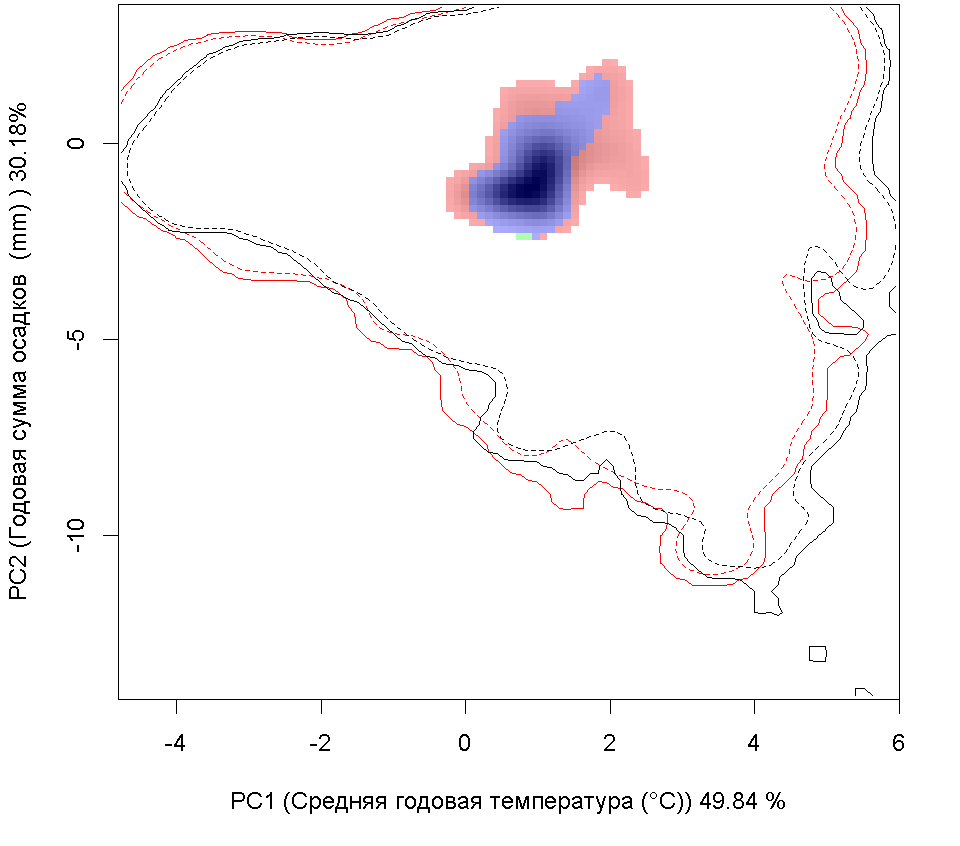
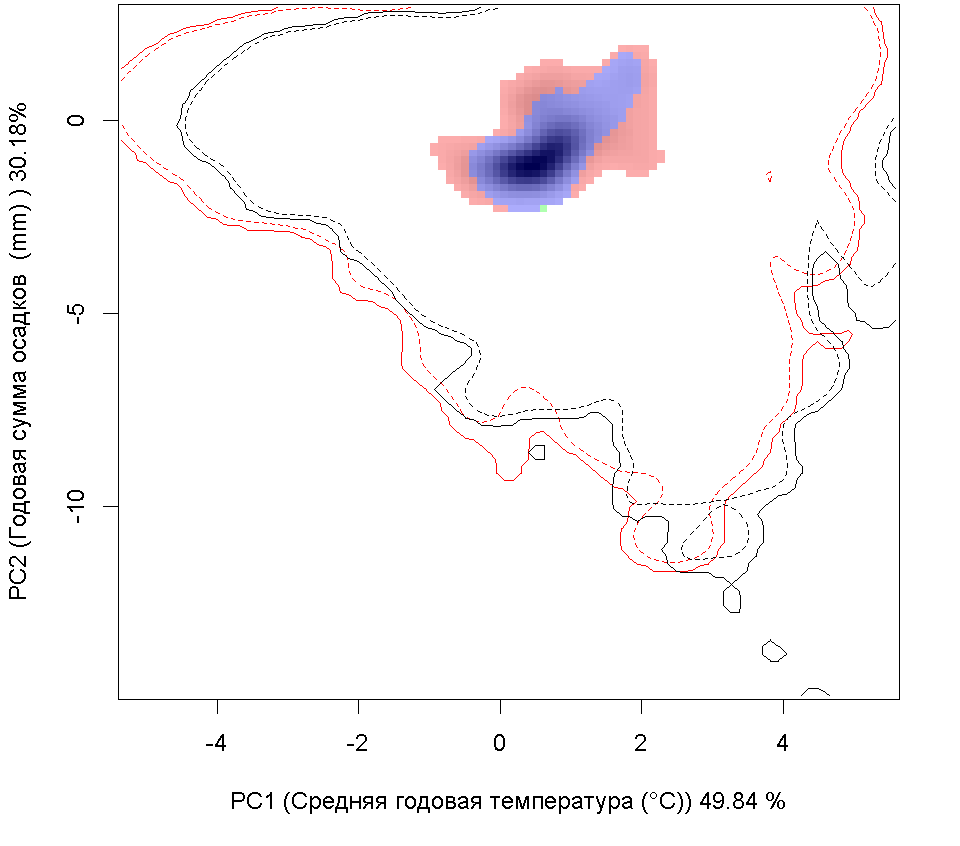
Сценарии изменения климата
|
|
RCP26
|
RCP45
|
|
 C C
|
 D D
|
|
RCP60
|
RCP85
|
|
 E E
|
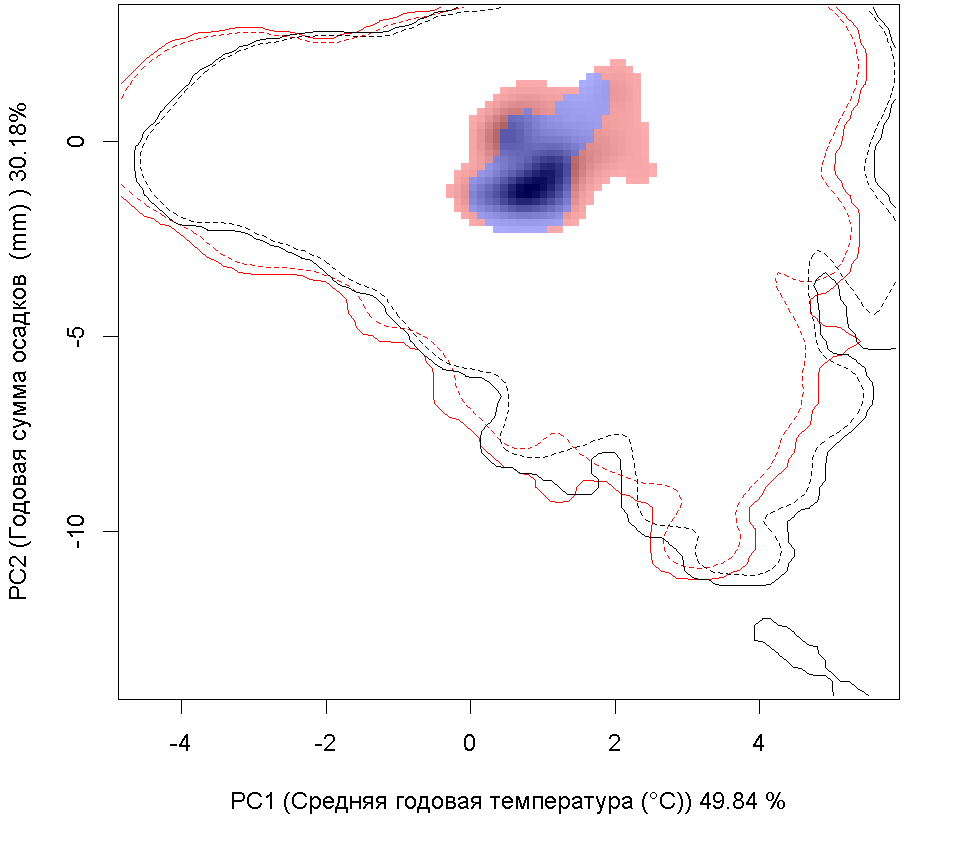 F F
|
| Графическое представление перекрывания ниш в условиях текущего климата и при сценариях его изменения - (C) RCP26; (D) RCP45; (E) RCP60; (F) RCP85. |
Литература
- Бабаназарова О.В. Кармайер Р., Сиделев С.И., Александрина Е.М. Сахарова Е.Г. Структура фитопланктона и содержание микроцистинов в высокоэвтрофном озере Неро // Вод. рес. — 2011. — 38(2). — С. 223—231.
- Бабаназарова О.В., Сиделев С.И., Фастнер Дж., 2014.Экспансия Сylindrospermopsis raciborskii (Nostocales, Cyanoprokaryota) в северные широты: вспышка развития в мелководном высокоэвтрофном оз. Неро (Россия). Альгология, 24 (4): 526-537.
- Бикбулатов Э.С., Бикбулатова Е.М., Литвинов А.С., Поддубный С.А. Гидрология и гидрохимия озера Неро. — Рыбинск: Рыбин. Дом печати, 2003. — 190 с.
- Кузьмин Г.В. Водоросли планктона Шекснинского и сопредельной акватории Рыбинского водохранилища // Биология, морфология и систематика водных организмов. — Л.: Наука, 1976. — C. 3—61.
- Протасов А.А., Силаева Л.П., Ярмошенко Л.П., Новоселова Т.Н., Примак А.Б., Савицкий А.Л. Гидробиологические исследования техно-экосистемы Запорожской АЭС // Гидробиол. журн. — 2013. — 49(2). — С. 78—94. Состояние экосистемы озера Неро в начале XXI века. — М.: Наука, 2008. — 406 с.
- Antunes J.T., Pedro N. Leao P.N., Vasconcelos V.M. 2015. Cylindrospermopsis raciborskii: review of the distribution, phylogeography, and ecophysiology of a global invasive species. Frontiers in Microbiology, 6:
- Babanazarova O.V. Phytoplankton structure, composition and production in saline waterbodies in the Lower Amu Dar’ya // Int. J. Salt Lake Res. — 1997. — 6. — P. 217—231.
- Babanazarova O.V., Lyashenko O.A. Inferring long-term changes in the physical-chemical environment of the shallow, enriched lake Nero from statistical and functional analyses of its phytoplankton // J. Plankt. Res. — 2007. — 29(9). — P. 747-756.
- Bernard C., Harvey M., Briand J.F., Bire R., Krys S., Fontaine J.J. Toxicological comparison of diverse Cylindrospermopsis raciborskii strains: evidence of liver damage caused by French C. raciborskii strain // Environ. Toxicol. — 2003. — 18(3). — P. 176—186.
- Bonilla S., Aubriot L., Soares M.C.S. et al. What drives the distribution of the bloomforming cyanobacteria Planktothrix agardhii and Cylindrispermopsis raiborskii? // Microbiol. Ecol. — 2012. — 79. — P. 594—607.
- Bourke ATC, Hawes RB, Neilson A et al. (1983) An outbreak of the hepatoenteritis (the Palm island mystery disease) possibly caused by algal intoxication. Toxicon (Suppl 3):45–48.
- Briand J.F., Robillot C., Quiblier-Lloberas C., Hubert J.F., Coute A., Bernard C. Environmental context of Cylindrospermopsis raciborskii (Cyanobacteria) blooms in a shallow pond in France // Water Res. — 2002. — 36(13). — P. 3183—3192.
- Briand J.-F., Leboulanger C., Humbert J.-F., Bernard C., Dufour P. Cylindrospermopsis raciborskii (Cyanobacteria) invasion at mid-latitudes: selection, wide physiological tolerance, or global warming ? // J. Phycol. — 2004. — 40(2). — P. 231—238.
- Chapman A.D., Schelske C.L. Recent appearance of Cylindrospermopsis (Cyanobacteria) in five hypereutrophic Florida lakes // J. Phycol. — 1997. — 33. — P. 191—195.
- Chonudomkul D., Yongmanitchai W., Theeragool G. et al. Morphology, genetic diversity, temperature tolerance and toxicity of Cylindrospermopsis raciborskii (Nostocales, Cyanobacteria) strains from Thailand and Japan // FEMS Microbiol. Ecol. — 2004. — 48. — P. 345—355.
- Dantas E.W., Bittencourt-Oliveira M.C., Moura A.N. Spatial-temporal variation in coiled and straight morphotypes of Cylindrospermopsis raciborskii (Wolosz.) Seenayya et Subba Raju (Cyanobacteria) // Acta Bot. Bras. — 2010. — 24(2). — P. 585—591.
- Druart J.C., Briand J.F. First record of Cylindrispermopsis raiborskii (Woloszynska) Seenayya et Subba Raju (Cyanobacteria) in a lotic system in France // Ann. Limnol. — 2002. — 38(4). — P. 339—342.
- Fastner J., Rucker J., Stuken A. et al. Occurrence of the cyanobacterial toxin cylindrospermopsin in Northeast Germany // Environ. Toxicol. — 2007. — 22. — P. 26—32.
- Hong Y., Steinman A., Biddanda B., Rediske R., Fahnenstiel G. Occurrence of the toxinproducing cyanobacterium Cylindrospermopsis raciborskii in Mona and Muskegon Lakes, Michigan // J. Great Lakes Res. — 2006. — 32. — P. 645—652.
- Istvanovics V., Shafik H.M., Presing M., Juhos S. Growth and phosphate uptake kinetics of the cyanobacterium, Cylindrospermopsis raciborskii (Cyanophyceae) in throughflow cultures // Fresh. Biol. — 2000. — 43(2). — P. 257—275.
- Komarek J., Horecka M. Taxonomic position of three planktonic blue-green algae from the genera Aphanizomenon and Cylindrospermopsis // Preslia Praha. — 1979. — 51. — P. 289—312.
- Moore D., O’Donohue M., Shaw G., Critchley C. Potential triggers for akinete differentiation in an Australian strain of the cyanobacterium Cylindrospermopsis raciborskii (AWT 205/1) // Hydrobiologia. — 2003. — 506-509. — P. 175—180.
- Ohtani I., Moore R.E., Runnegar M.T.C. Cylindrospermopsin: a potent hepatotoxin from the blue-green alga Cylindrospermopsis raciborskii // J. Amer. Chem. Soc. — 1992. — 114. — P. 7941—7942.
- Padisak J. Cylindrospermopsis raciborskii (Woloszynska) Seenayya et Subba Raju, an expanding, highly adaptive cyanobacterium: worldwide distribution and review of its ecology // Arch. Hydrobiol. (Suppl.) — 1997. — 107. — P. 563—593.
- Padisak J. Estimation of minimum sedimentary inoculum (akinete) pool of Cylindrospermopsis raciborskii: a morphology and life-cycle based method // Hydrobiologia. — 2003. — 502. — P. 389–394.
- Padisak J., Crosesetti L.O., Naselli-Flores L. Use and misuse in the application of the phytoplankton functional classification: a critical review with updates // Ibid. — 2009. — 621. — P. 1–19.
- Reynolds C.S. Phytoplankton periodicity: the interactions of form, function and environmental variability // Freshwat. Biol. — 1984. — 14. — P. 111–142.
- Reynolds C.S., Huszar V., Kruk C., Naselli-Flores L., Melo S. Towards a functional classification of the freshwater phytoplankton // J. Plankt. Res. — 2002. — 24(5). — P. 417–428.
- Saker M.L., Neilan B.A. Varied diazotrophies, morphologies, and toxicities of genetically similar isolates of Cylindrospermopsis raciborskii (Nostocales, Cyanophyceae) from Northern Australia // Appl. and Environ. Microbiol. — 2001. — 67(4). — P. 1839–1845.
- Shafik H.M., Voros L., Sprober P., Presing M., Kovacs A.W. Some special morphological features of Cylindrospermopsin raciborskii in batch and continuous cultures // Hydrobiologia. — 2003. — 506-509. — P. 163–167.
- Sidelev S.I., Fastner J., Dittmann-Thunemann E., Babanazarova O.V., Zubishina A.A. First report on the presence of cylindrospermopsin-producing blue-green algae (Cyanobacteria) in the Russian waters // Mat. II(X) Int. bot. conf. of young sci. in St.-Petersburg (Saint-Petersburg, 11–16 Nov. 2012). — St.Pb., 2012. — P. 40. Singh R.N. Seasonal variants of Anabaenopsis raciborskii Wolosz. // Hydrobiologia. — 1962. — 20. — P. 87–91.
|