Представлена версия документа, предназначенная для печати
Вы можете загрузить эту страницу на сайте
Erigeron annuus (L.) Pers., 1807 |
Систематическое положение:
Царство — растения, Plantae. Отдел — сосудистые, Tracheophyta. Класс — двудольные, Magnoliopsida. Порядок — астроцветные, Asterales. Семейство – астровые, Asteraceae. Вид — мелколепестник однолетний, Erigeron annuus

Русское и английское названия. Мелколепестник однолетний Annual fleabane
Синонимы. Daisy fleabane; eastern daisy fleabane; tall fleabane; Aster annuus L., A. stenactis E.H.L. Krause, Erigeron annuus var. discoideus (Victorin & J. Rousseau) Cronquist, E. strigosus var. septentrionalis (Fernald & Wiegand) Fernald, Doronicum bellidiflorum Schrank, Phalacroloma annuum (L.) Dumort., Ph. annuum subsp. annuum, Ph. septentrionale (Fernald & Wiegand) Tzvelev, Stenactis annua (L.) Less., S. septentrionalis (Fernald & Wiegand) Holub.
Нативный ареал. Северная Америка: восток США, южная Канада.
Современный ареал (мировой и конкретнее в России). Вторичный ареал включает Европу, Азию (Анатолию, Китай, Корею, Японию, Непал, Дальний Восток, Индию), Северную и Центральную Америку, Новую Зеландию (Черная книга…, 2016). В Европе отмечен в 32 странах, в 15 из них натурализовался (Lambdon et al., 2008). Наибольшее распространение характерно для умеренных областей Европы.
В России мелколепестник однолетний распространен в основном на территории европейской части, где встречается в 34 областях, в 27 из них натурализовался (Морозова и др., 2013), на Северном Кавказе и Дальнем Востоке. Области наибольшего распространения лежат в широколиственнолесной и лесостепной областях. На Дальнем Востоке отмечен для Сахалина, Курильских островов и юга Приморского края (Баркалов, 1992). В 2009 г. обнаружен на территории Центрального Сибирского Ботанического сада в Новосибирске (в Западной Сибири) как сорное растение (Лащинский и др., 2010).
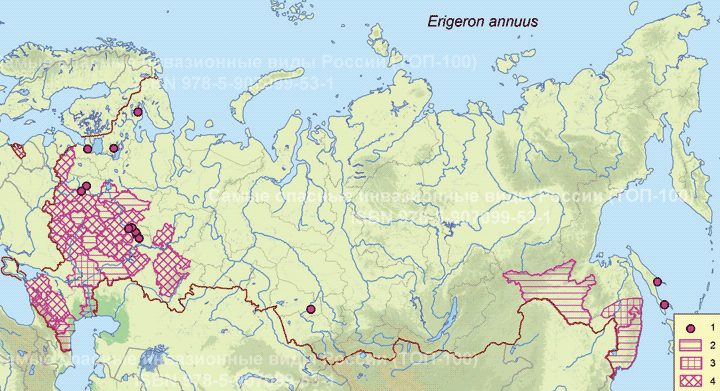
Пути(коридоры) и векторы (способы) интродукции. Выращивался в ботанических садах с XVII в. (впервые упомянут из ботанического сада в Париже в 1636 г.), а затем как декоративное растение с начала XIX в. Первые находки в Европе — в Дании в 1810 г., в Бельгии в 1813 г., в Нидерландах в 1826 г, в Швейцарии в 1828 г. (Frey, 2003). В России отмечен позже, первые находки известны из Московской области: в 1900 г. А.Н. Петунниковым по сборам К.Л. Гейдена из парка Горенки Балашихинского района (Игнатов и др., 1900), однако первый сбор был в 1902 г. (MW, Д. Сырейщиков, Бол. Мытищи).
Первоначально выращивался в качестве декоративного растения, дальнейшее расселение обусловлено свойствами самого вида, для которого основной способ распространения анемохория. Н.М. Решетниковой (2016) высказано предположение о наличии у вида зоохорного распространения семян. Может быть занесен с семенами других растений (Лащинский и др., 2010).
Местообитание. В естественном ареале растет в основном на открытых нарушенных местообитаниях, обочинах дорог, полях, лугах, мусорных местах. Во вторичном ареале встречается на сорных местах, залежах, у дорог, на лугах, по сухим склонам и поймам рек, на лесных полянах. Доминирует в сообществах начальных сукцессионных стадий зарастания залежей (Stratton, 1992; Клюев, 2013). Предпочитает легкие почвы (Майоров и др., 2012).
Некоторые особенности биологии.Однолетний апомиктический вид. Свойства вида, обеспечивающие его инвазионный «успех»: 1) высокий генетический полиморфизм, несмотря на наличие апомиксиса, что можно объяснить редким перекрестным опылением; 2) высокая семенная продуктивность: одно растение образует от 10 тыс. до 100 тыс. семян (Stratton, 1991; Frey, 2003; Виноградова и др., 2009).
Влияние вида (на другие виды, экосистемы включая лесную и агроценозы, здоровье человека). Сорное растение. Обладает аллелопатическими свойствами, может ингибировать прорастание семян некоторых видов.
Контроль численности. Вид внесен в Черную книгу флоры Средней России (Виноградова и др., 2009). Специальных мер борьбы не разработано.
Автор
Морозова О.В.|
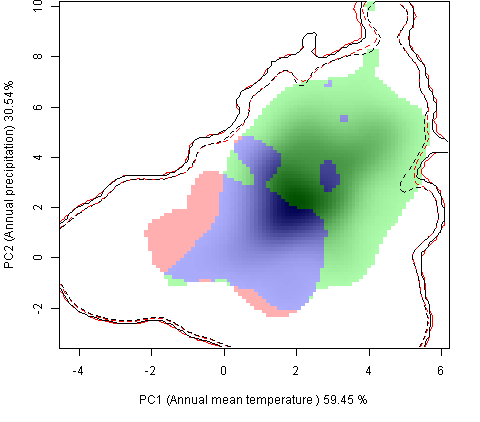
Графическое представление перекрывания ниш нативной и инвазионной частей ареалов, а также при сценариях изменения климата |
|
|
Текущий климат |
|
|
Нативная часть |
Инвазионная часть |
|
|
|
| Графическое представление перекрывания ниш нативной (A) и инвазионной (B) частей ареалов вида, где сиреневый цвет – зона стабильности, розовый цвет – зона расширения, зеленый цвет – зона «неиспользования». Сплошные и пунктирные линии показывают 100% и 90% области доступной среды в нативной (черные линии) и инвазионной (красные линии) частях ареалов, которые использовались для анализа перекрывания ниш. | |
|
Сценарии изменения климата |
|
|
RCP26 |
RCP45 |
|
|
|
|
RCP60 |
RCP85 |
|
|
|
| Графическое представление перекрывания ниш в условиях текущего климата и при сценариях его изменения - (C) RCP26; (D) RCP45; (E) RCP60; (F) RCP85. | |
Литература
| |
кабинетом «Биоинформатики и моделирования биологических процессов» ИПЭЭ РАН в рамках выполнения гранта РНФ № 16-14-10323. | |
| © 2004-. Кабинет "Биоинформатики и моделирования биологических процессов" ИПЭЭ РАН | |
 A
A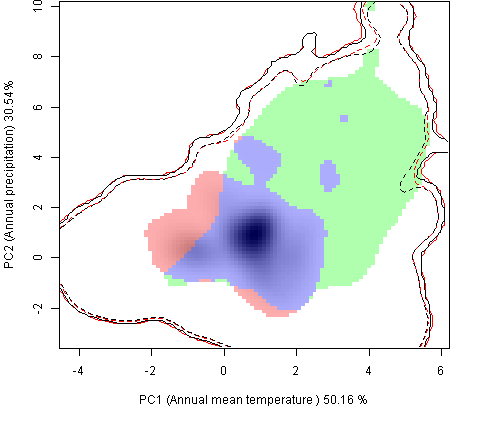 B
B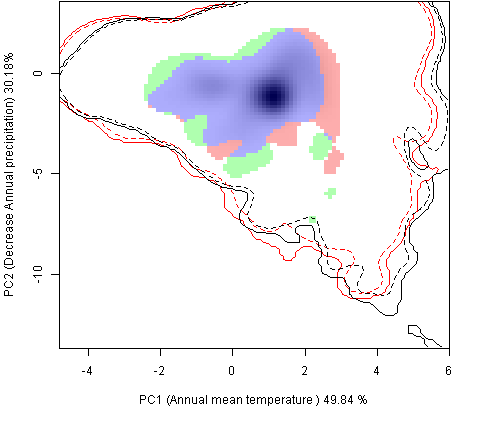 C
C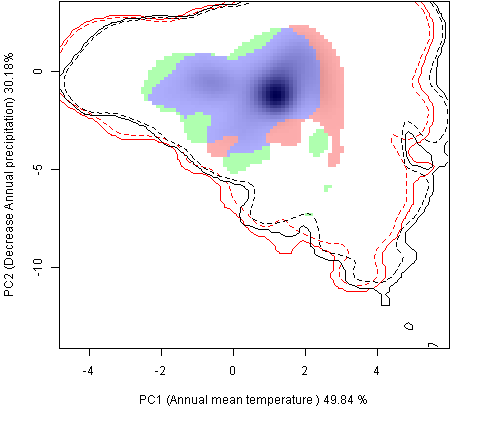 D
D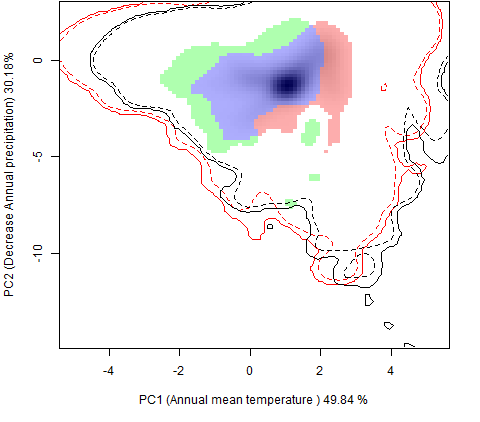 E
E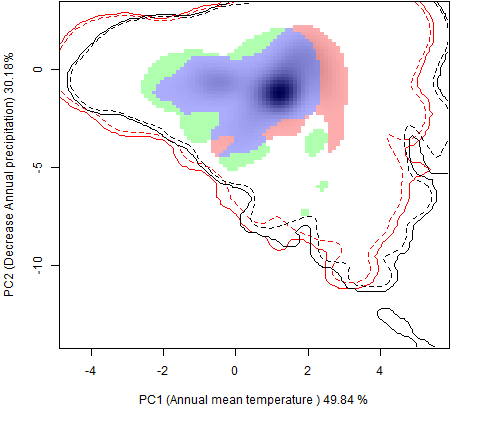 F
F