Представлена версия документа, предназначенная для печати
Вы можете загрузить эту страницу на сайте
Mus musculus Linnaeus, 1758 |
Систематическое положение (Kingdom, Phylum, Class, Order, Family, Species):
Animalia » Chordata » Mammalia » Rodentia » Muridae » Mus musculus L.
Животные Animalia » Хордовые Chordata » Млекопитающие Mammalia » Грызуны Rodentia » Мышиные Muridae » Домовая мышь Mus musculus L.

Русское и английское названия. Домовая мышь, House Mouse.
Синонимы. Имеет несколько внутривидовых форм, которые некоторые авторы рассматривают на уровне видов. Шире всех распространена M. m. domesticus: Зап. Европа, в Азии – страны Средиземноморья, Африка (без Мадагаскара), С. и Ю. Америки, Австралия. В России в подавляющем большинстве регионов M. m. musculus, распространены также ее гибриды с формами: wagneri (юг Европейской России), gansuensis (Тува, Забайкалье), castaneus (Дальний Восток), реже с domesticus и bactrianus. В некоторых зарубежных публикациях M. m. musculus или M. musculus s.s. называют: Eastern house mouse или Eurasian house mice Устоявшихся русских названий этих форм – нет. Ниже, когда это касается домовых мышей в любой части Земли и не оговаривается особо, речь идет о синантропных формах домовых мышей (Mus musculus s.l.), а для территории России о Mus m. musculus и её гибридах.
Нативный ареал. Юго-Западная Азия
Современный ареал (мировой и конкретнее в России). Расселилась по всем материкам, кроме Антарктиды, и островам всех океанов. В России распространена почти повсеместно, за исключением высокогорий Кавказа и малонаселенных частей севера и северо-востока. Далеко на север проникает только с человеком. Показано, что российская часть ареала относительно хорошо совпадает с освоенной территорией (плотность населения – более 1 чел. на 1 кв. км), установлена высокая степень совпадения густоты пунктов регистрации домовой мыши с пахотными угодьями (Кучерук, 1994). Инвазионная часть ареала существенно превышает нативную часть.
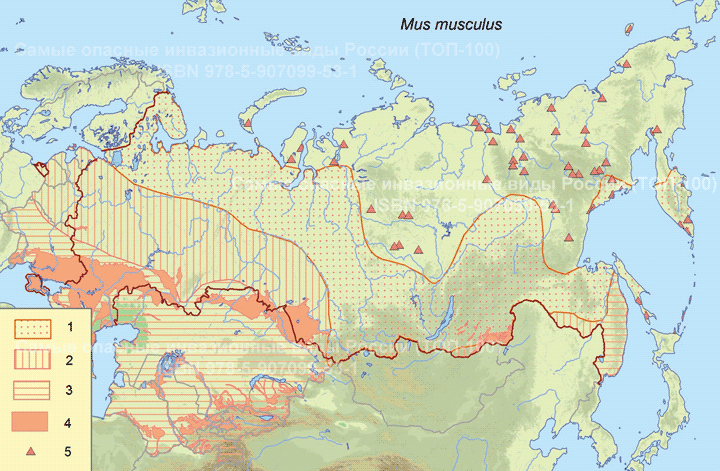
Пути(коридоры) и векторы (способы) интродукции. Домовые мыши - древнейшие и постоянные спутники человека. Их расселение и обширный ареал, прежде всего, связаны с синантропностью и агрофильностью вида, т.е. обитанием в созданных человеком местообитаниях – населенных пунктах и полях зерновых. От первых этапов появления земледелия до наших дней человек, расселяясь сам и перевозя пищевые продукты и, прежде всего, зерно, развозит вместе с собой домовых мышей. История формирования ареала сложная, далеко нерешенная задача. Современные молекулярно-генетические исследования подтверждают древность расселения, формирование нескольких основных генетических линий и наличие зон гибридизации, которые могут занимать обширные пространства (Cucchi et al., 2005; Searle et al., 2009; Спиридонова и др., 2008; 2011; Suzuki et al., 2013; Спиридонова, 2014; Gray et al., 2014; Jing et al., 2014; Jones, Searle, 2015; Liu et al., 2015; Мальцев и др., 2016 и др.).
Начало заселения домовыми мышами юга современной территории России уходит в далекое прошлое освоения этой территории племенами древних людей и их переходом к жизни в настоящих жилищах и земледелию (около 10 тыс. лет до н.э.) (Лавренченко, 1994). На плодородных землях аридных территорий в полной мере могла проявляться и развиваться экологическая пластичность вида, сочетающая адаптации к обитанию в дикой природе, к синантропии и к агрофилии. К районам древнего обитания домовых мышей на территории современной России, по-видимому, следует отнести освоенные древними людьми участки степных и околоводных пустынных местообитаний юга европейской части России, юга Сибири и Дальнего Востока. По мере развития Российского государства, освоения и расширения его земель происходило дальнейшее расселение домовой мыши, для которого существенное значение имели транспортные потоки. В.В. Кучерук (1994) показал, что основным средством расселения домовых мышей служили перевозки зерна водным транспортом. Для eвропейской части СССР это были реки бассейнов Черного, Каспийского и Белого морей. Для Сибири, за исключением ее южной части, основными путями расселения служили и продолжают служить судоходные реки. В бассейнах Печоры, Оби, Енисея и Лены развоз мышей осуществлялся в двух направлениях: с юга (центральных частей материка) на север и с севера (от морских побережий) на юг. По рекам Яна, Индигирка, Колыма, Анадырь – только вглубь материка. В Якутии и Магаданской области большую роль для расселения домовых мышей сыграли автомобильные магистрали: Беркатит-Якутск, Ленск-Мирный, Якутск-Вилюйск-Кемпендяй, Якутск-Магадан, Магадан-Усть Нера, Магадан-Омсукчан. Мыши могут развозиться и по железным дорогам, но в России они успели заселить трассы будущих рельсовых магистралей раньше, чем началось их строительство (первая железная дорога в России была открыта в 1837 г.). Не исключены перевозки авиатранспортом. Проникновение домовых мышей в лесные и тундровые части территории России шло вместе с формированием сети сельских и городских населенных пунктов, где мыши обитали в жилищах, выходя из них летом в центральных частях России на посевы или (реже) в природные биотопы. На значительной части территории России домовые мыши – древние инвайдеры, однако север Сибири и северо-восток России заселяли во второй половине ХХ в. (Бобров и др., 2008; Хляп, Варшавский, 2009), что связано с бурным экономическим развитием этих регионов и ростом транспортных потоков. Доказательством продолжающегося современного расселения домовых мышей служит обнаружение M m. domesticus на северо-востоке России, где до 2000 г. таких находок не было (Yonekawa et al., 2003). Другим фактом широкого переселения домовых мышей вместе с человеком служат обширные зоны гибридизации, не типичные для мелких млекопитающих, расселяющихся естественными путями (Котенкова, 2002; Kotenkova, 2004; Фрисман и др., 2010; Спиридонова и др., 2011 и др.)
Местообитание. Основные места обитания: населенные пункты, поля зерновых, в степях — тростниковые заросли. Домовая мышь теплолюбива. Не встречается в высокогорьях. В России повсеместно обитает в городских и сельских населенных пунктах, регионально – на полях зерновых и в природных биотопах. По возможности домовых мышей обитать вне жилищ различают 3 зоны: южная, центральная и северная (Тупикова, 1947; Хляп, Варшавский, 2009). В южной зоне (аридные территории юга Европейской России и Приморье) домовые мыши могут круглогодично жить на пахотных землях, а также в природных местообитаниях. Среди последних наиболее благоприятны тростниковые заросли вдоль водоемов. Новыми местами концентрации домовых мышей становятся берега созданных в XX в. оросительных каналов и водохранилищ. Кружево ареала и плотность поселений домовых мышей в южной зоне в значительной степени определяются площадью и конфигурацией полей зерновых и способа уборки урожая. В результате распашки общий запас численности этих грызунов здесь увеличился по сравнению с доагрикультурным временем в сотни раз (Тупикова и др., 2000; Неронов и др., 2001; Хляп, Варшавский, 2010). В центральной зоне мыши могут покидать жилье человека только летом, выселяясь на 1-2 км от жилья на приусадебные земли, поля зерновых, в заросли бурьяна, единично в кустарники и леса. В северной зоне (северная тайга и тундры) домовая мышь обитает только в жилищах человека. Однако чумы и землянки оленеводов-кочевников для нее не пригодны. Граница ареала и плотность его кружева в центральной и северной зонах тесно связаны с динамикой людских поселений, и территория, заселенная домовыми мышами, постоянно меняет свои очертания, т.к. в жилых постройках домовые мыши появляются, а с ликвидацией жилья – исчезают (Кучерук, 1994).
Некоторые особенности биологии, обуславливающие расселение вида и влияние на экосистемы, другие виды и на человека. Характеризуется высокой подвижностью, легко преодолевая малопригодные для нее биотопы (Никитина, 1958; Хляп и др., 1986). Характерно смешанное питание. Преобладают семена, роль которых возрастает к осени, велика доля зеленых кормов, насекомых, реже поедает ягоды (Окулова и др., 2011). Предпочитает влажные местообитания, склонна к обитанию на полях и в населенных пунктах. Нередко достигает высокой численности (Плятер-Плохоцкий, 1936; Шкилев, 1960; Кулик, 1971; Тихонова и др., 1992; Окулова и др., 2011; 2012). Сезонный пик размножения в Центральном Черноземье приходится на июнь-август, а в Приамурье – на август. Средняя величина выводка в Центральном Черноземье варьирует от 4,7 до 7,8. В Тамбовской области и в Приамурье отмечено подснежное зимнее размножение (Окулова и др., 2012). Полевой мыши свойственна высокая экологическая пластичность, которая позволяет адаптироваться к антропогенно нарушенной среде с помощью разнообразных популяционных, в том числе поведенческих, механизмов (Москвитина, Сучкова, 1994; Агулова и др., 2008). Все эти экологические характеристики обуславливают адаптации полевых мышей к антропогенным биотопам и их широкое самостоятельное расселение, особенно вдоль ручьев, по уремам рек и с/х землям. Заселяет сельскохозяйственную продукцию (сено, контейнеры с овощами и т.п.), с которой может перевозиться человеком на дальние расстояния, например, завезена в Северное Приохотье (Переверзева и др., 2016).
Некоторые особенности биологии, обуславливающие расселение вида и влияние на экосистемы, другие виды и на человека.Домовые мыши - древнейшие и постоянные спутники человека. Расселение и широкий ареал этого вида, прежде всего, связаны с его синантропностью, т.е. обитанием на большей части ареала в населенных пунктах, и с агрофильностью , т.е. с обитанием летом на полях зерновых на юге постоянно, а севернее сезонно. Н.В. Тупикова (1947) пришла к выводу, что домовую мышь как синантропа отличает отсутствие специфических приспособлений и экологическая пластичность. Эти зверьки обитают в жилище человека и даже предпочитать их другим биотопам т.к.: а) всеядны, а отсюда не испытывают недостатка в корме; б) не имеют узкой специализации в типе передвижений, хорошо лазают, бегают, прыгают; развитый грызущий аппарат и небольшие размеры дают возможность проникать во все части зданий и переезжать с товарами; в) не требовательны к обстановке гнездования; г) имеют лабильный ритм активности, который позволяет приспособиться к расписанию людей в заселенных мышами помещениях; д) характеризуются хорошо развитой терморегуляцией и поведенческими особенностями, позволяющими теплолюбивой мыши приспособиться к разнообразным температурным условиям. По мнению Е.В. Котенковой (2000) разгадка феномена синантропии кроется в уникальном сочетании эколого-этологических и физиологических характеристик домовых мышей. Им присущи свойства тропических видов (нечувствительность к фотопериоду), видов аридных территорий (физиологические приспособления к минимальной потери влаги и ее постоянному дефициту), высокая стрессоустойчивость и уникальный набор эколого-этологических адаптаций, включающих: лабильность пространственно-этологической структуры; наличие в популяции лабильного резерва особей, способных к размножению; высокую исследовательскую активность и др. Становление синантропного образа жизни имело весомое значение в процессе эволюции домовых мышей (Котенкова, Мунтяну, 2007).
Другая черта биологии домовых мышей – агрофилия – на первых этапах эволюции вида неразрывно сочеталась с синантропией, т.к. жилища человека и возделываемые им поля располагались поблизости. Позже синантропные и агрофильные популяции разделились в пространстве. Интенсивная распашка земель стала существенной причиной изменения ареала, роста численности домовых мышей и повышения их ценотической роли (Тупикова и др., 2000; Неронов и др., 2001; Хляп, Варшавский, 2010). Развитие городов и сети поселений человека позволили мышам обитать в холодных широтах.
Влияние вида (на другие виды, экосистемы включая лесную и агроценозы, здоровье человека). Домовые мыши относятся к особой группе чужеродных видов, т.к. основной вред наносят человеку (Хляп и др., 2010). Они поедают и загрязняют продукты питания., при высокой численности наносят ощутимый урон своей грызущей деятельностью. В домах могут заражать людей лимфоцитарным хориоменингитом и псевдотуберкулезом. Обнаружены антитела к Yersinia enterocolitica. В регионах, где домовые мыши могут постоянно или периодически жить в природе, они включаются в циркуляцию туляремийного микроба, а на юге возбудителей чумы и др. инфекций (Кулик, 1979; Левченко и др., 2014). При миграции в населенные пункты могут служить причиной заноса возбудителей в жилища человека. В учебных заведениях г. Омска находили домовых мышей, зараженных возбудителем токсоплазмоза (Сидоров, Путин, 2010). Показано, что, поселяясь в жилых постройках, домовые мыши могут препятствовать вселению туда рыжих полевок (Myodes glareolus Schreber, 1780), что уменьшает возможность заражения людей геморрагической лихорадкой с почечным синдромом (Краснов, Хохлова, 1989).
Контроль численности. Контроль численности домовых мышей, как и других синантропных грызунов, имеет глубокую историю. Чаще применяют физические (отлов) и химические (яды-антикоагулянты) методы ограничения численности. При химическом методе истребления мышей необходимо учитывать, что домовые мыши относительно устойчивы к ядам, и концентрация действующих веществ в приманках от мышей должна быть выше, чем в отравленных приманках от крыс. Показано, что антикоагулянты II поколения более эффективны в отношении домовых мышей. Хорошие результаты дает также применение препаратов, содержащих витамин D. Имеются некоторые различия в приемах проведения истребительных мероприятий. Мыши не любят долго кормиться в одном месте, поэтому можно раскладывать маленькие порции отравленных приманок, но их должно быть много. Из-за высокой устойчивости к ядам нередко отлов мышей бывает эффективней, чем применение отравленных приманок. Современные подходы к изъятию грызунов (особенно в природных местообитаниях островов) предусматривают проведение предварительных исследований, включая диагностику видовой принадлежности грызунов (Campbell et al., 2015).
|
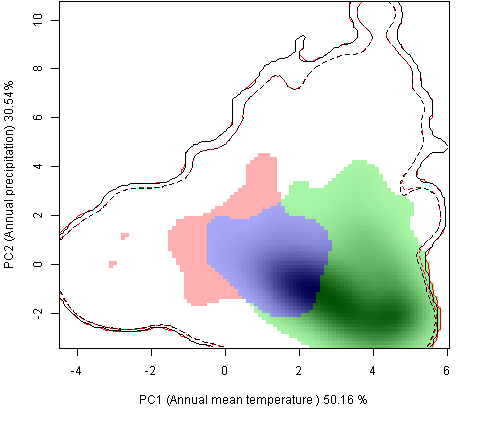
Графическое представление перекрывания ниш нативной и инвазионной частей ареалов, а также при сценариях изменения климата |
|
|
Текущий климат |
|
|
Нативная часть |
Инвазионная часть |
|
|
|
| Графическое представление перекрывания ниш нативной (A) и инвазионной (B) частей ареалов вида, где сиреневый цвет – зона стабильности, розовый цвет – зона расширения, зеленый цвет – зона «неиспользования». Сплошные и пунктирные линии показывают 100% и 90% области доступной среды в нативной (черные линии) и инвазионной (красные линии) частях ареалов, которые использовались для анализа перекрывания ниш. | |
|
Сценарии изменения климата |
|
|
RCP26 |
RCP45 |
|
|
|
|
RCP60 |
RCP85 |
|
|
|
| Графическое представление перекрывания ниш в условиях текущего климата и при сценариях его изменения - (C) RCP26; (D) RCP45; (E) RCP60; (F) RCP85. | |
Литература
| |
кабинетом «Биоинформатики и моделирования биологических процессов» ИПЭЭ РАН в рамках выполнения гранта РНФ № 16-14-10323. | |
| © 2004-. Кабинет "Биоинформатики и моделирования биологических процессов" ИПЭЭ РАН | |
 A
A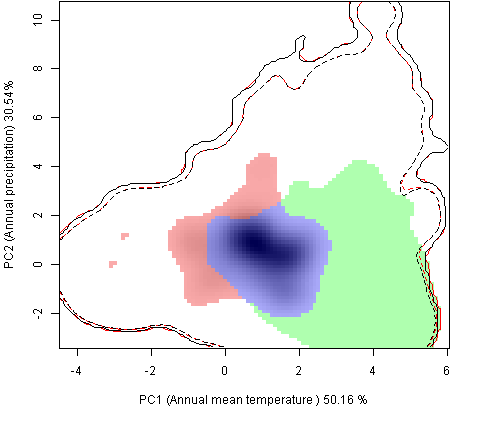 B
B C
C D
D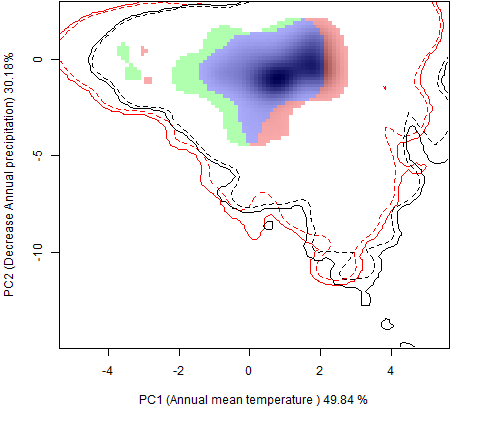 E
E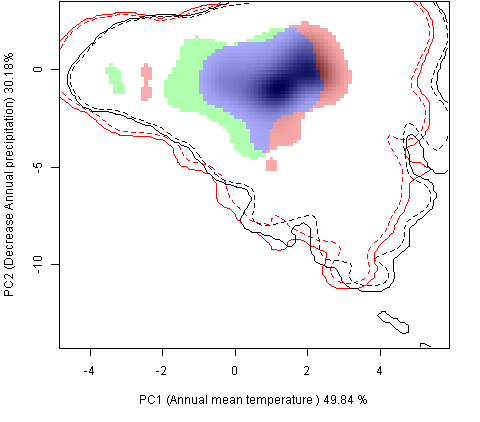 F
F