Представлена версия документа, предназначенная для печати
Вы можете загрузить эту страницу на сайте
Ophiostoma novo-ulmi |
Систематическое положение (Kingdom, Phylum, Class, Order, Family, Species):
Fungi » Ascomycota » Sordariomycetes » Ophiostomatales » Ophiostomataceae » Ophiostoma novo-ulmi

Русское и английское названия. Dutch elm disease, Офиостома вязовая
Синонимы. Цератоцистис вязовый, Ceratocystis ulmi, Pesotum ulmi (Anamorph)
Нативный ареал. Родиной патогенного для ильмовых деревьев грибка Ophiostoma novo-ulmi считают Юго-Восточную Азию. Включает два подвида O. novo-ulmi subsp. americana и О. novo-ulmi subsp. novo-ulmi. Оба подвида имеют Азиатское происхождение.
Современный ареал (мировой и конкретнее в России). Возбудитель голландской болезни ильмовых (или графиоз ильмовых) впервые занесен в Голландию, откуда распространился по всей Европе и проник в Северную Америку. Продолжает распространяться в северных районах Европы, особенно в Шотландии. Потепление климата также может способствовать его расширению в северном направлении. Массовое поражение этим грибом вяза в Москве и Московской обл. отмечено в конце 1990-х гг. Позднее он проник в более северные регионы. В частности, в начале ХХ в. эпифитотия голландской болезни вязов отмечена в озеленительных насаждениях Санкт-Петербурга. Известен из Вологодской, Ленинградской, Московской, Самарской, Саратовской, Ростовской и Астраханской областей, Республики Калмыкии. В С.-Петербурге в 2008-2015 гг. проводили мониторинг голландской болезни вязов, результаты которого отражены на сайте: http://grafioz.myopencity.org/map.html

Пути (коридоры) и векторы (способы) интродукции. Известно, что вязовые леса Европы и России погибли в результате двух эпифитотий. Первая - в западноевропейских странах в 1920-е гг. и в России в 1930-е гг.- была вызвана инвазией в леса возбудителя Ophiostoma ulmi. В Европе болезнь впервые появилась в 1917—1919 годах в Нидерландах. Причем тогда Голландия потеряла до 70% вязовых насаждений. Предполагают, что грибок мог быть завезён с корзинами из вязовых прутьев, в которых перевозили вещи китайские рабочие. В 1919—1929 годах болезнь вязов распространилась по Европе. В 1930—1933 годах с грузами брёвен была занесена в США через порты Атлантики и Мексиканского залива. В 1936 году голландская болезнь достигла западных регионов СССР и юго-западной Азии, а к 1940 году эпидемия в Европе практически прекратилась, но продолжалась в Америке, распространившись по восточным и центральным штатам. В 1940 году болезнь была обнаружена на востоке Канады. Урон среди насаждений вяза американского достигал до 15% в год. Примерно с 1940-х гг. в лесах Европы (Украина, Румыния) началась вторая волна голландской болезни вязов, которую связывают с «возвращением» грибка из Америки. Исследования показали, что грибок, как на американском, так и европейском континенте существенно эволюционировал. Новая патогенная форма была описана в 1991 г. как самостоятельный вид - Ophiostoma novo-ulmi (Brasier , 1991, 1995, 2001). Болезнь быстро распространилась в некоторых Европейских странах. Особенно пострадала Великобритания, где вяз считается одним из национальных символов. Во многих регионах насаждения были полностью уничтожены. К XXI веку голландская болезнь вязов оказалась широко распространённой в центральных и южных регионах европейской части России. Страны, где растут преимущественно мелколистные вязы, например, Австрия, пострадали незначительно. Переносчиками являются жуки-короеды: заболонник большой ильмовый (Scolytus scolytus), заболонник струйчатый (Scolytus multistriatus), реже заболонник пигмей (Scolytus pygmaeus) (в Европе), американский ильмовый короед (Hylurgopinus rufipes) (в Америке). Могут участвовать в распространении и листогрызущие насекомые — ильмовый листоед (Xanthogaleruca luteola) и другие. Гриб образует бесполые спороношения и плодовые тела в галереях, прогрызенных жуками и личинками. Как конидии, так и аскоспоры покрыты слизью и легко прилипают к телу жука, а время спороношения (весной) совпадает со временем лёта жуков. Подсыхая, споры могут также разноситься ветром и распространяться вместе с заражённой древесиной. Заражение в этом случае происходит при попадании спор на свежие повреждения.
Местообитание. Болезнь может распространяться в естественных лесах, лесных культурах, защитных лесополосах и городских посадках. В чистых вязовых насаждениях риск эпифитотии выше, чем в смешанных.
Влияние вида (на другие виды, экосистемы включая лесную и агроценозы, здоровье человека). Голландская болезнь ильмовых пород деревьев относится к категории наиболее опасных сосудистых болезней деревьев. Это инфекционное усыхание кроны, которое в большинстве случаев приводит к гибели дерева. Грибок офиостома интенсивно выделяет токсин увядания — цератоульмин, что приводит к быстрой гибели деревьев. Заболевание деревьев может носить острый характер (вся крона усыхает в течение одного сезона) или хронический характер (крона дерева усыхает ветка за веткой в течение нескольких лет). При хронической форме болезни усыхание деревьев происходит в течение 8-10 лет. У крупных старых деревьев с хронической формой заболевания уже в середине мая листва заметно мельче, чем у здоровых деревьев - особенно это выражено в верхней части кроны. На протяжении всего лета такие деревья выделяются ажурной кроной с единичными усохшими ветвями. Острая форма болезни вызывает усыхание деревьев за один вегетационный период, месяц или даже несколько дней. В настоящее время микологи выделяют три штамма вызывающих голландскую болезнь вязов: Ophiostoma ulmi, Ophiostoma novo-ulmi и Ophiostoma himal-ulmi. Считается, что гибель вязовых лесов С. Америки, Европы и России вызвана поражением деревьев патогенными офиостомами. В результате его патогенного воздействия только в Великобритании погибло около 28 млн. вязов, в Северной Америке – сотни миллионов вязов. В настоящее время большинстве европейских стран находиться в пост эпидемиальной ситуации.
|
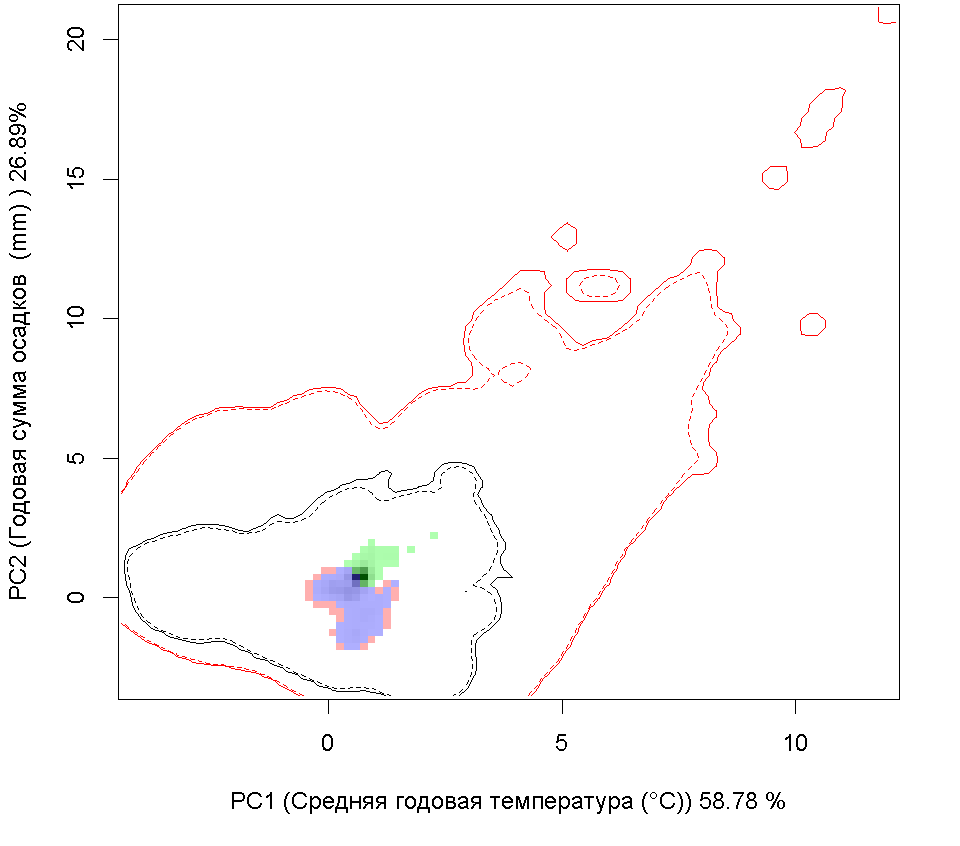
Графическое представление перекрывания ниш нативной и инвазионной частей ареалов, а также при сценариях изменения климата |
|
|
Текущий климат |
|
|
Нативная часть |
Инвазионная часть |
|
|
|
| Графическое представление перекрывания ниш нативной (A) и инвазионной (B) частей ареалов вида, где сиреневый цвет – зона стабильности, розовый цвет – зона расширения, зеленый цвет – зона «неиспользования». Сплошные и пунктирные линии показывают 100% и 90% области доступной среды в нативной (черные линии) и инвазионной (красные линии) частях ареалов, которые использовались для анализа перекрывания ниш. | |
|
Сценарии изменения климата |
|
|
RCP26 |
RCP45 |
|
|
|
|
RCP60 |
RCP85 |
|
|
|
| Графическое представление перекрывания ниш в условиях текущего климата и при сценариях его изменения - (C) RCP26; (D) RCP45; (E) RCP60; (F) RCP85. | |
Литература
Другие ссылки
| |
кабинетом «Биоинформатики и моделирования биологических процессов» ИПЭЭ РАН в рамках выполнения гранта РНФ № 16-14-10323. | |
| © 2004-. Кабинет "Биоинформатики и моделирования биологических процессов" ИПЭЭ РАН | |
 A
A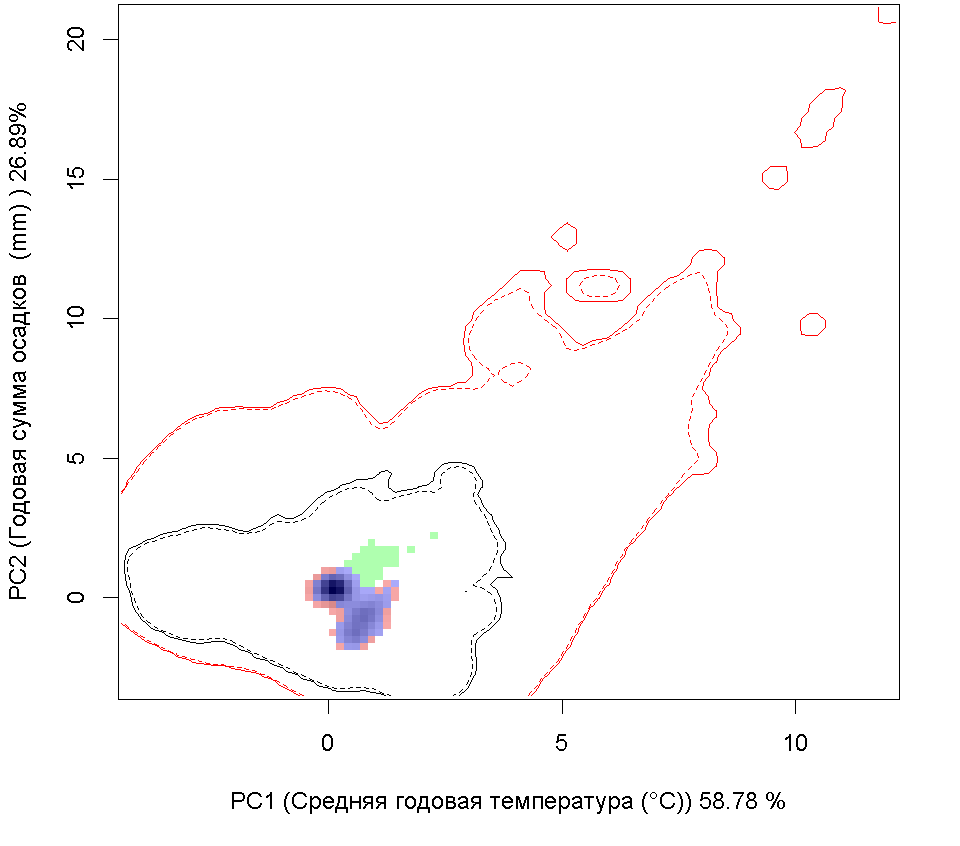 B
B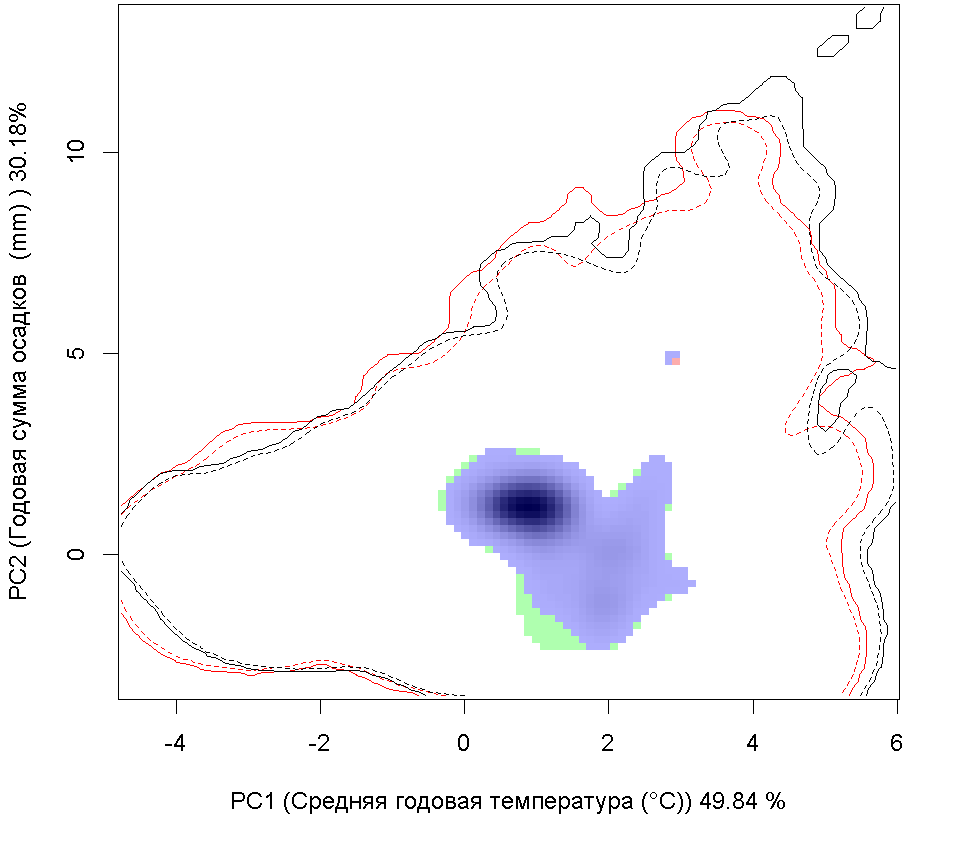 C
C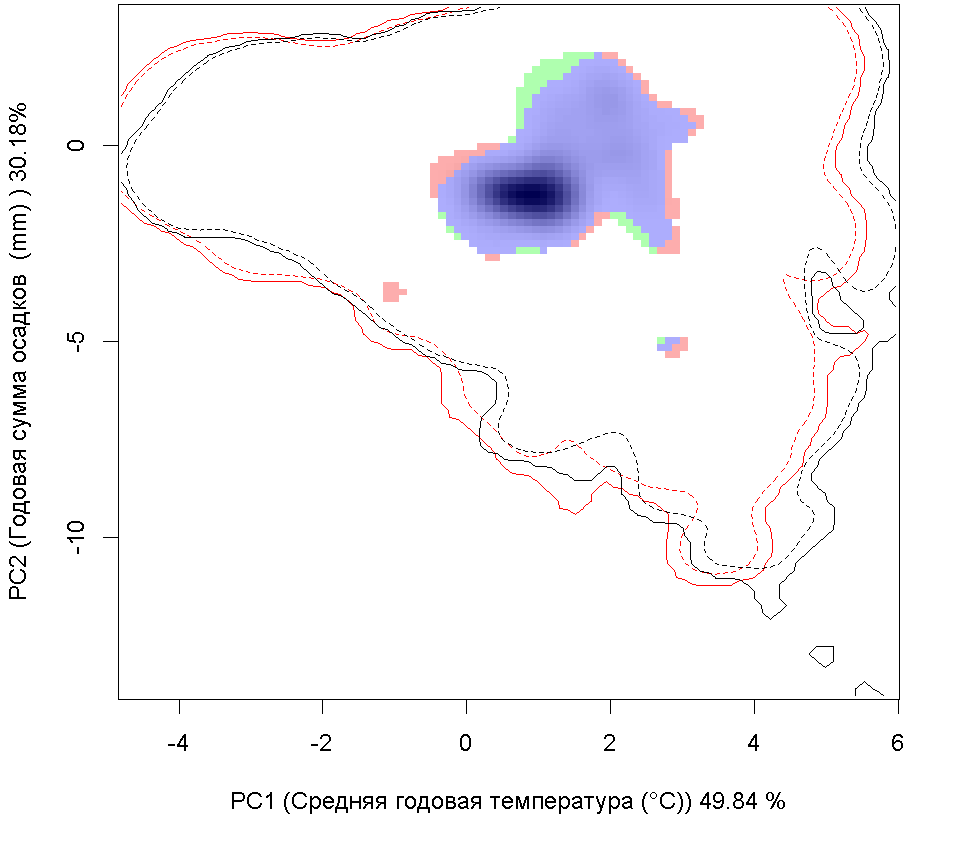 D
D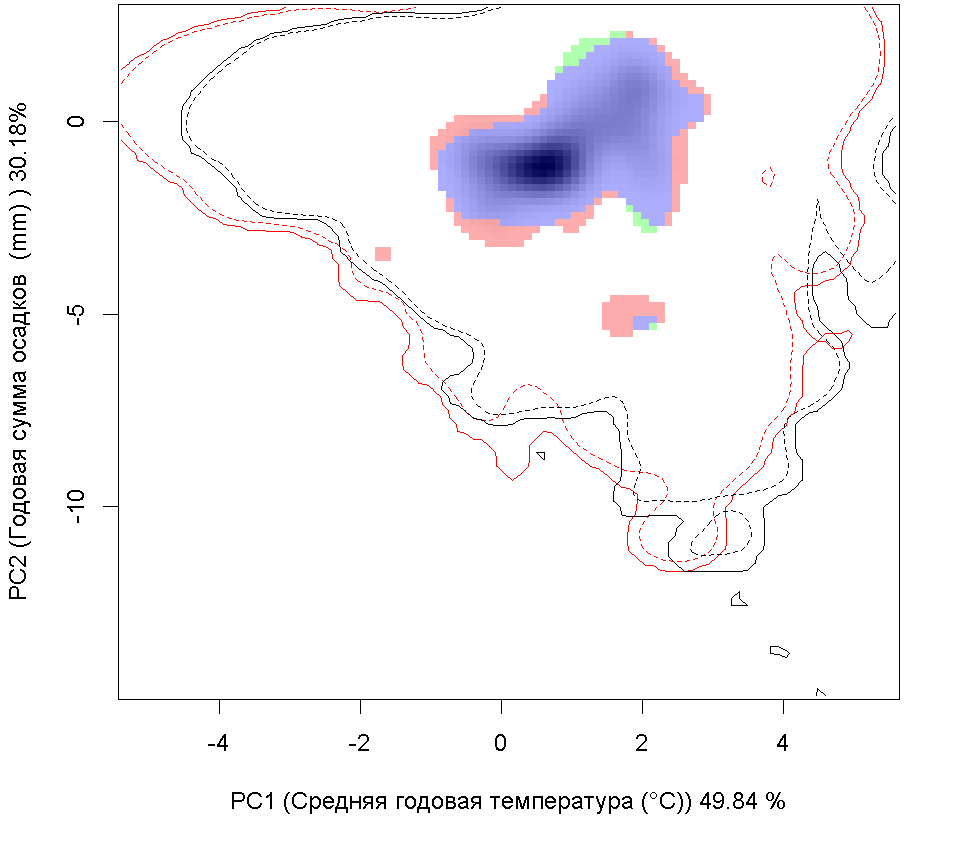 E
E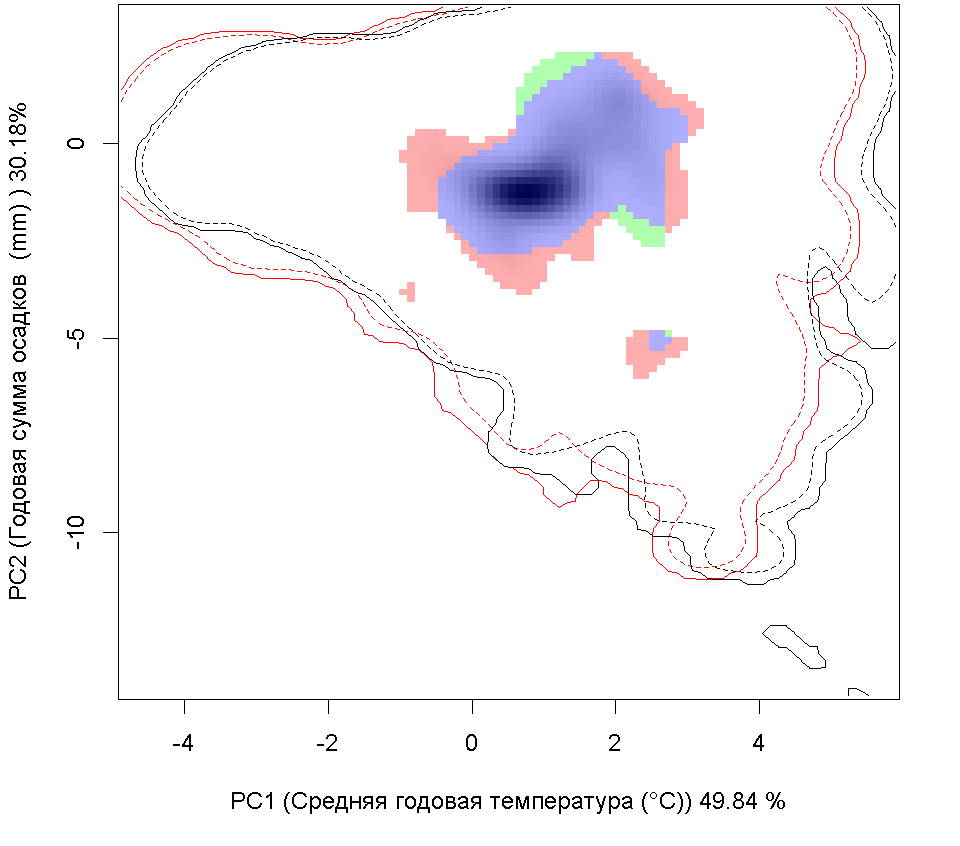 F
F