Представлена версия документа, предназначенная для печати
Вы можете загрузить эту страницу на сайте
Monocorophium acherusicum (Costa, 1853) |
Систематическое положение (Kingdom, Phylum, Class, Order, Family, Species):
Царство – животные. Animalia. Тип – членистоногие, Arthropoda. Класс – высшие раки, Malacostraca. Отряд – бокоплавы, Amphipoda. Семейство – корофииды, Corophiidae. Вид – Monocorophium acherusicum
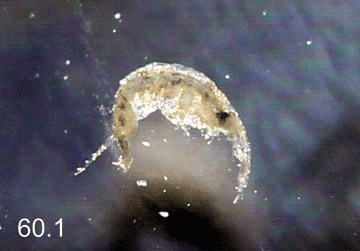
Русское и английское названия. нет
Синонимы. Названий на русском и английском языках нет Podocerus cylindricus Say, 1818; Audouinia acherusica Costa in Hope, 1851; Corophium acherusicum Costa, 1853; Corophium cylindricum (Say, 1818) (в Smith, 1873 согласно Stebbing, 1914); Corophium contractum Stimpson, 1855 (согласно Stebbing, 1914); Corophium bonelli H. Milne Edwards, 1830 и Corophium crassicorne Bruzelius, 1859 (в Совинский, 1880 согласно Cranford, 1937), Corophium bonnellii (H. Milne Edwards, 1830) (в Barnard, 1932 согласно Cranford, 1937); Corophium quadriceps Dana, 1852 (согласно Stebbing, 1914); Corophium crassicorne Bruzelius, 1859 (в Hoek, 1879 согласно Cranford, 1937).
Нативный ареал. По последним данным предполагается, что нативным ареалом этой амфиподы является атлантическое побережье Северной Америки, включая находки в Кубе и Пуэрто-Рико (Stebbing, 1906; Shoemaker, 1934). Криптогенным вид считается на атлантическом побережье Европы и Африки, а в остальных местах обитания – инвазионным (Albano et al., 2018). Вид был впервые упомянут в 1851 г. (Costa in Hope, 1851) и затем описан в 1853 г. по материалам, собранным в озере Фузаро, отделенного от Тирренского моря узкой полосой суши (провинция Неаполь, Италия, бассейн Средиземного моря) (Costa, 1853). Однако предполагается, что первые находки этого виды были сделаны на атлантическом побережье Северной Америки, которые были в то время идентифицированы как Podocerus cylindricus Say, 1818 (Shoemaker, 1934).
Современный ареал (мировой и конкретнее в России). Вид – космополит. Распространен в умеренных, субтропических и тропических регионах. Обитает в бассейнах Атлантического, Тихого и Индийского океанов. На атлантическом побережье Северной Америки M. acherusicum отмечен в Канаде, США и Мексике. В Европе на атлантическом побережье он встречается на территории от Испании до Шотландии. Зарегистрирован у берегов Дании в проливах Большой и Малый Бельт, дальше в Балтийское море вид не заходит. M. acherusicum населяет бассейн Средиземного моря, в том числе Черное море (Турция, Россия, Болгария). Обнаружен на островах Атлантического океана – Азорских, Канарских, Фолклендских, Куба и Пуэрто-Рико. У атлантического побережья Африки он обитает от Марокко до Сенегала, в Кот-д’Ивуаре, Намибии и ЮАР, у атлантического побережья Южной Америки – в Венесуэле, Бразилии и Аргентине. В пределах Индийского океана встречается в ЮАР, Танзании, Пакистане, Индии, Австралии и на островах Крусадай и Маврикий. В Тихом океане зарегистрирован у берегов Австралии, Новой Зеландии, Чили, Перу, Таиланда, Китая, Кореи, Японии, России, Канады, США, Мексики и Гавайских островов. Находки этого вида на Аляске и в других районах на широтах выше 60° с.ш. в настоящее время считаются сомнительными.
В России он отмечен у побережья Крыма. На востоке страны известен у берегов Японского моря, включая залив Петра Великого, и на восточном шельфе Сахалина (подробней ниже).
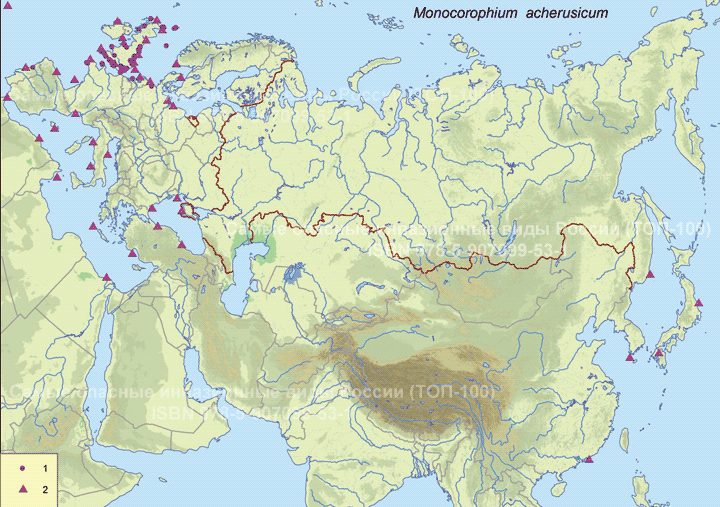
Пути(коридоры) и векторы (способы) интродукции. M. acherusicum относится к группе амфипод, строящих трубки (tube-building amphipod). Распространение вида осуществляется в основном балластными водами (Albano et al., 2018).
Первые находки в криптогенной части ареала были сделаны на атлантическом побережье Евразии и Средиземном море, во второй половины XIX в. и первой половине ХХ в. В Атлантическом океане у берегов Европы вид был обнаружен у берегов: Нидерландов – 1879 г. (Hoek, 1879), Франции – 1887 г. (Chevreux, 1900), Испании и Португалии (Chevreux, 1911), Англия (Шотлнадия) – 1913 г. (Walker, 1914) и в Средиземном море: Италии – 1851 г. (Costa in Hope, 1851), России – 1878 г. (Совинский, 1880), Турции – 1892-1893 гг. (Совинский, 1887), Алжире и Тунисе – 1908 (Chevreux, 1908), Египте и Франции – 1911 (Chevreux, 1911). Формирование инвазионной части ареала шло по разным направлениям и, судя по всему, завозы были неоднократными. Первая регистрация в инвазионной части ареала установлена в Тихом океане и датируется серединой XIX в.: Япония – 1855 г. (Stimpson, 1855), затем в Китае (Гонг Конг) – 1906 г. (Stebbing, 1906). В это время вид также проникает в Суэцкий канал (Египет), где его обнаруживают у г. Суэц в 1909 г. (Walker, 1909).
Во второй половине XX в. и начале XXI в. освоение прибрежных акватории Евразии продолжилось. Первые находки M. acherusicum в новых регионах датируются следующим образом. Тихий океан у берегов Евразии: Россия – 1970 г. (Зевина и др., 1975), Южная Корея – 1981 г. (Hong, 1983), Таиланд – 1994 г. (Angsupanich, Kuwabara, 1999). Атлантический океан у берегов Европы: Ирландия – 2007 г. (Daniels et al., 2009), Германия – 2009 г. (Schückel et al., 2010), Дания (проливы Большой и Малый Бельт) – 2012 г. (HELCOM, 2012) и в Средиземном море: Греция – 1976 г. (Stefanidou, Voultsiadou-Koukoura, 1995), Болгария – 2012 г. (Uzunova, 2012). Индийский океан: Индия – 1970 (Sivaprakasam, 1970), Пакистан – 1976 г. (Ahmad, 1976 цит. по: Kazmi, Khatoon, 2016). Дальнейшее распространение M. acherusicum возможно вдоль тихоокеанского побережья Евразии на север от Японского моря, а также вероятно проникновение в Азовское море.
Впервые о находке M. acherusicum в России в районе Карадага у Керченского полуострова (Черное море) упоминают Н.М. Милославская и В.Л. Паули (1931). Однако есть мнение (Cranford, 1937 и др.), что еще раньше (в 1878 г.) эта амфипода была поймана в Севастопольской бухте В. Совинским (1880). С последней трети XX в. вплоть до настоящего времени находки у побережья Крыма не редкость (Маккавеева, 1972; Евстигнеева и др., 2015 и др.). В Японском море у берегов России вид был впервые обнаружен в 1970 г. (Зевина и др., 1975). После этого он активно распространился в акватории залива Петра Великого (Будникова, 2001; Белогурова, Звягинцев, 2006 и др.) и найден на восточном шельфе Сахалина (Будникова, Савко, 2002). Авторы (А.А. Гусев, Д.О. Гусева) обнаружили эту амфиподу в бухте Большой Камень, Уссурийский залив, Залив Петра Великого в 2017 г.
Местообитание. M. acherusicum населяет эстуарные экосистемы и встречается в обрастаниях как на мягких грунтах, так и на твердых (Bousfield, 1973). в приливо-отливных районах всех континентов, кроме Антарктиды. Населяет глубины от 0 до 20 м, иногда их можно обнаружить на глубинах до 90 м (Chevreux, 1911; Tasso et al., 2018 и др.). Живут при температуре воды от 0 °С до 30 °С и солености 5-38‰, однако предпочитают соленость выше 20‰ (Lee et al., 2003; Peterson, Vaysierres, 2010 и др.).
Особенности биологии. M. acherusicum – это небольшие амфиподы с размерами тела до 4-6 мм (Bousfield, 1973; Onbe, 1966). Продолжительность жизни до 12 месяцев (Joeng et al., 2006). Эти амфиподы строят трубки, в которых живут, выделяя похожие на шелк нити, к которым прикрепляются частицы детрита и осадка (Barnard et al. 1988). Трубки прикреплены к грунту или любым иным поверхностям, включая вертикальные: утесы, камни, сваи, понтоны, поплавки, буи, веревки, сети, корпуса судов, ракушки, высшая водная растительность, гидроиды, водоросли и губки (Barnard, 1958; Crawford, 1937; Onbe, 1966 и др.). M. acherusicum ведут малоподвижный образ жизни в дневное время, но способны плавать и переходить к планктонному образу жизни в ночное время, после сильного шторма или усиленного речного стока (Grabe, 1996). Амфиподы выходят из своего укрытия в поисках пищи и для спаривания. Они довольно активно мигрируют, так как одними из первых появляются на новых субстратах (Barnard, 1958). Питаются фитопланктоном и органическим детритом и, возможно, способны поедать донные микроводоросли с поверхности грунта и пастись на нитчатых эпифитных водорослях, которые развиваются на морских водорослях (Bousfield, 1973). Основными хищниками являются рыбы (Amundred et al., 2015; Onbe, 1966), возможно, крабы и креветки.
M. acherusicum – раздельнополые животные (Bousfield, 1973). Самки, как правило, крупнее самцов и могут достигать длины 6 мм против 4,5 мм у самцов (Onbe, 1966; Bousfield, 1973). В умеренном климате размножение сезонное с мая по сентябрь (Bousfield, 1973) или круглогодичное в южных районах, но более частое и начинается с меньших размеров тела самки (летом 2,2 мм, зимой 3,0 мм). Количество яиц и эмбрионов в выводковой камере самок с одной стороны связано с размером тела, 2-4 яйца у самых маленьких самок и 60-70 яиц у крупных самок длиной 6 мм, с другой, с высокой индивидуальной вариабельностью. Молодь, выходящая из выводковой камеры самок, по своему строению полностью похожа на взрослые особи и способна к самостоятельной жизни. На процесс размножение оказывает значительное влияние температура воды. Так при высоких температурах летом процесс размножения снижается, а в холодный сезон года в умеренной зоне – прекращается (Onbe, 1966).
Влияние вида (на другие виды, экосистемы включая лесную и агроценозы, здоровье человека). M. acherusicum широко распространенный вид-обрастатель. Создавая высокие плотности поселений на искусственных субстратах, он разрушительно влияет на трубопроводы, корпуса судов, доковые сооружения и объекты аквакультуры (Crawford, 1937; Barnard, 1958; Onbe, 1966). Он является важным потребителем фитопланктона, бентических и эпифитных микроводорослей, а с другой стороны – значимым объектом питания прибрежных рыб. Является экосистемным инженером, создает среду обитания, которую используют другие организмы. В пределах плотных поселений корофиумов затруднено проникновение в толщу грунта представителей инфауны (Barnard, 1958). В Японии обрастание M. acherusicum негативно повлияло на культуры водорослей и устриц (Onbe, 1966). Высокая плотность корофиид может оказать воздействие на стабильность донных отложений и увеличить скорость их эрозии в устьях рек (Talman et al., 1999).
Контроль численности. Поскольку вид распространяется с балластными водами и на корпусах судов, то для сдерживания распространения корофиид необходимо вести мониторинг в портах Японского, Охотского, Черного и Азовского морей, контроль и обезвреживание балластных вод, а также сбрасывать балластные воды в открытой части океана. Необходимо контролировать прибрежный туризм, чтобы предотвратить случайные заносы на свободные от M. acherusicum прибрежные территории. Среди важных аспектов управления и контроля – создание карт размещения M. acherusicum и разработка специальной программы мониторинга. В настоящее время методы биологической и химической борьбы с данным видом не разработаны.
Авторы. Гусев А.А., Гусева Д.О., Фенева И.Ю.
|
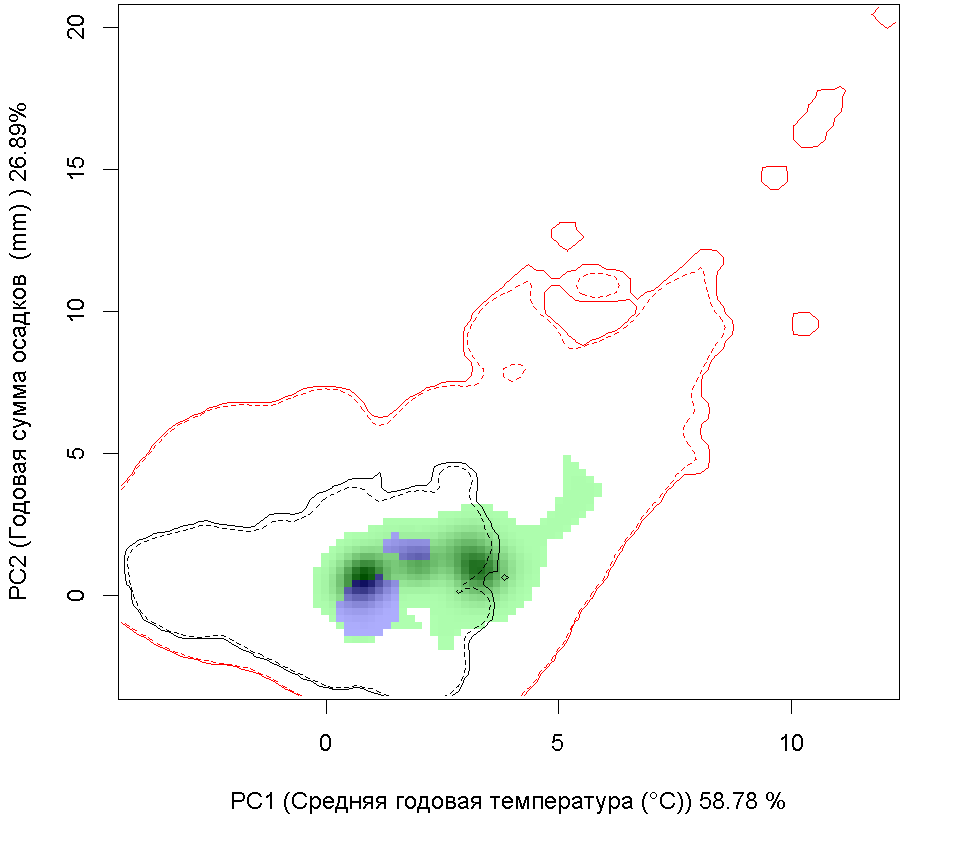
Графическое представление перекрывания ниш нативной и инвазионной частей ареалов, а также при сценариях изменения климата |
|
|
Текущий климат |
|
|
Нативная часть |
Инвазионная часть |
|
|
|
| Графическое представление перекрывания ниш нативной (A) и инвазионной (B) частей ареалов вида, где сиреневый цвет – зона стабильности, розовый цвет – зона расширения, зеленый цвет – зона «неиспользования». Сплошные и пунктирные линии показывают 100% и 90% области доступной среды в нативной (черные линии) и инвазионной (красные линии) частях ареалов, которые использовались для анализа перекрывания ниш. | |
|
Сценарии изменения климата |
|
|
RCP26 |
RCP45 |
|
|
|
|
RCP60 |
RCP85 |
|
|
|
| Графическое представление перекрывания ниш в условиях текущего климата и при сценариях его изменения - (C) RCP26; (D) RCP45; (E) RCP60; (F) RCP85. | |
Литература.
| |
кабинетом «Биоинформатики и моделирования биологических процессов» ИПЭЭ РАН в рамках выполнения гранта РНФ № 16-14-10323. | |
| © 2004-. Кабинет "Биоинформатики и моделирования биологических процессов" ИПЭЭ РАН | |
 A
A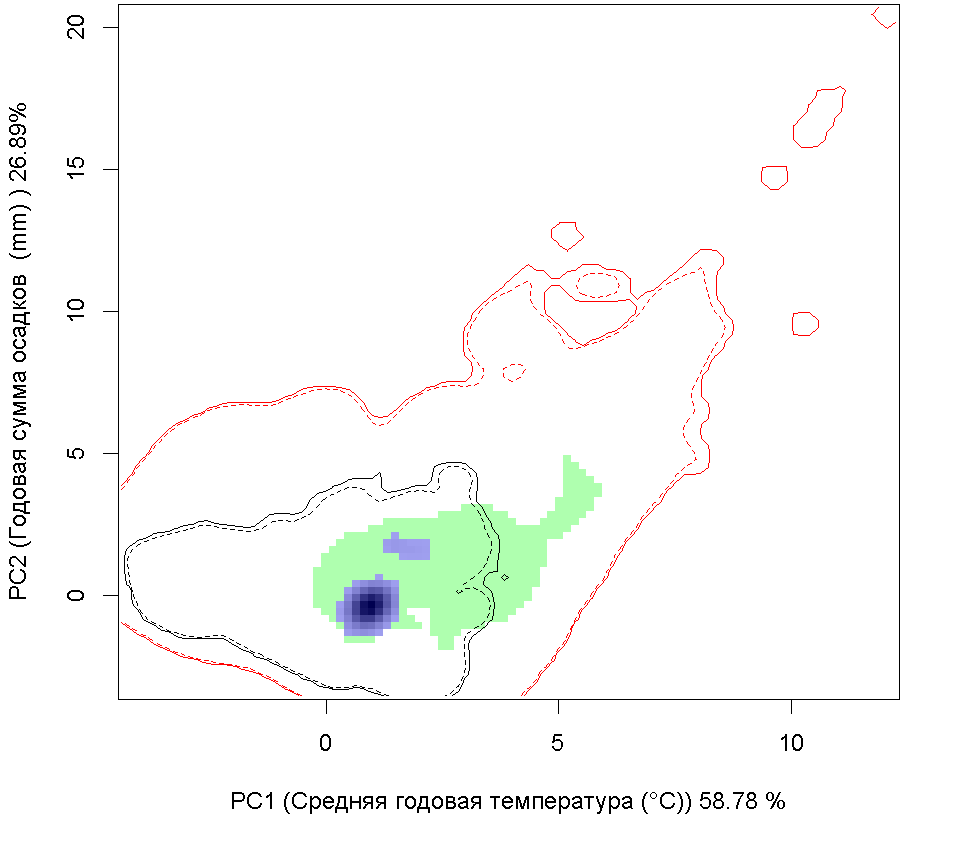 B
B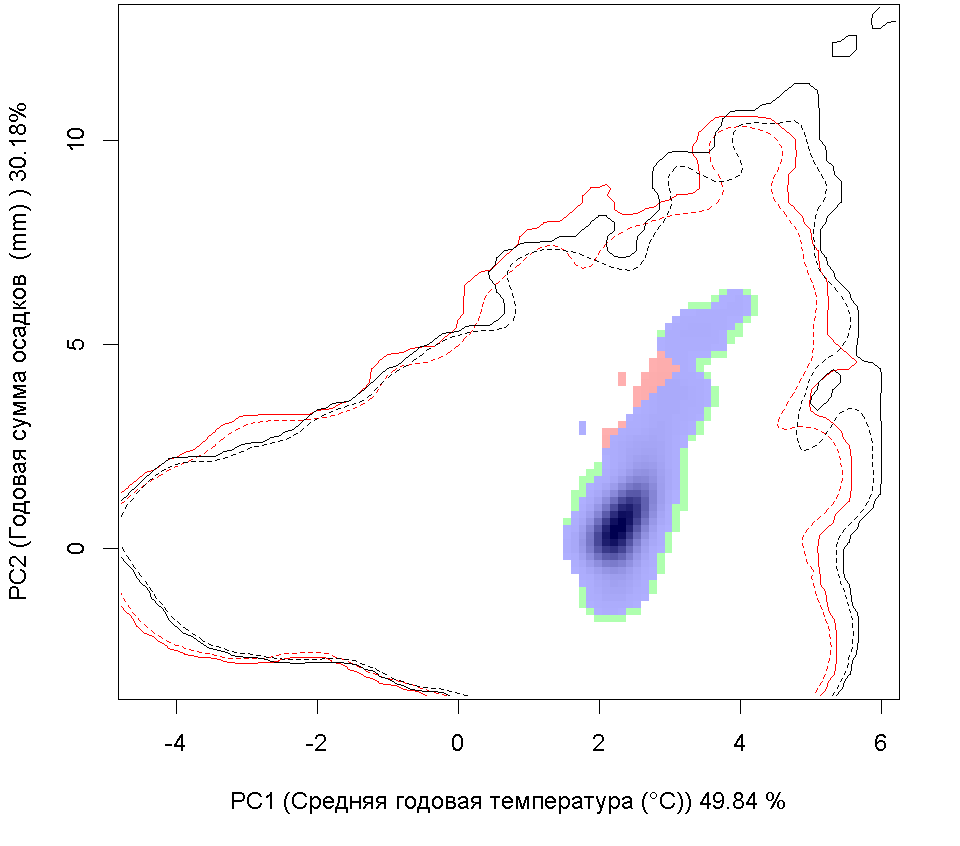 C
C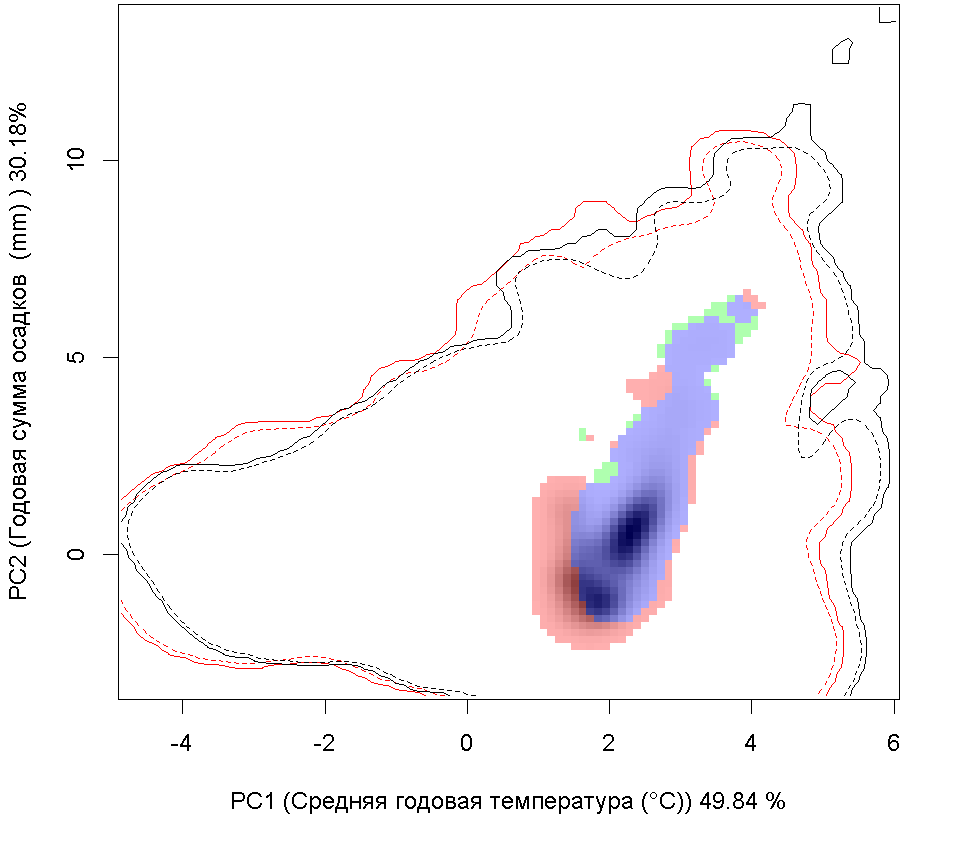 D
D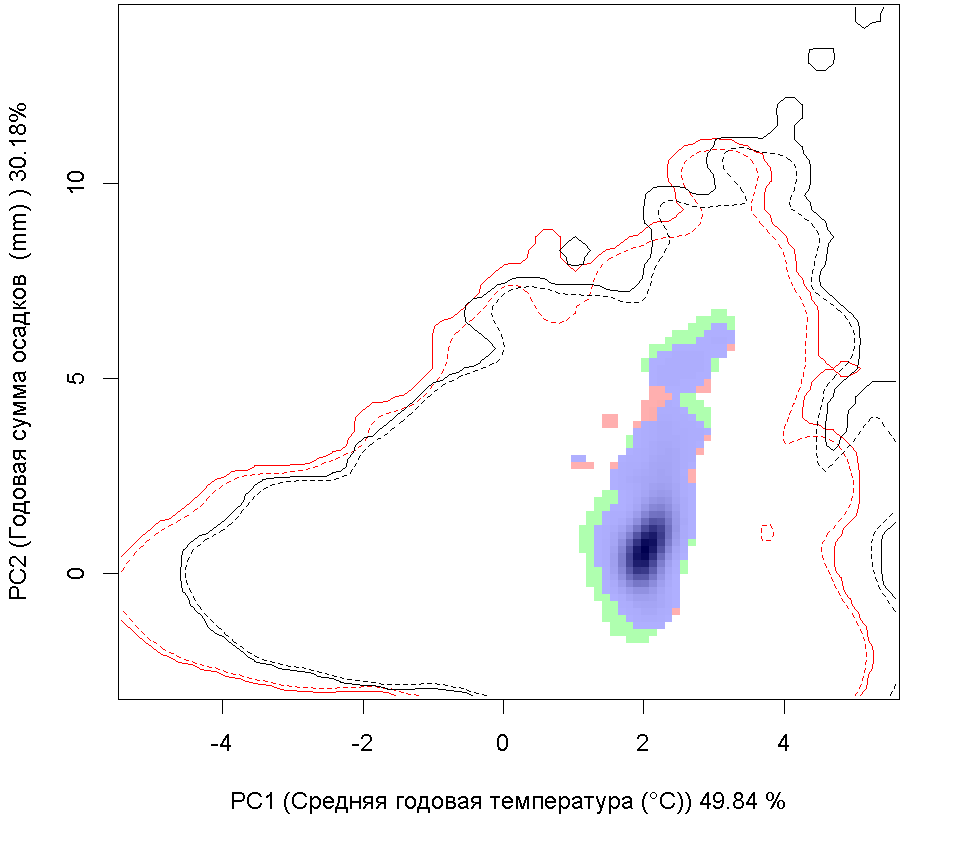 E
E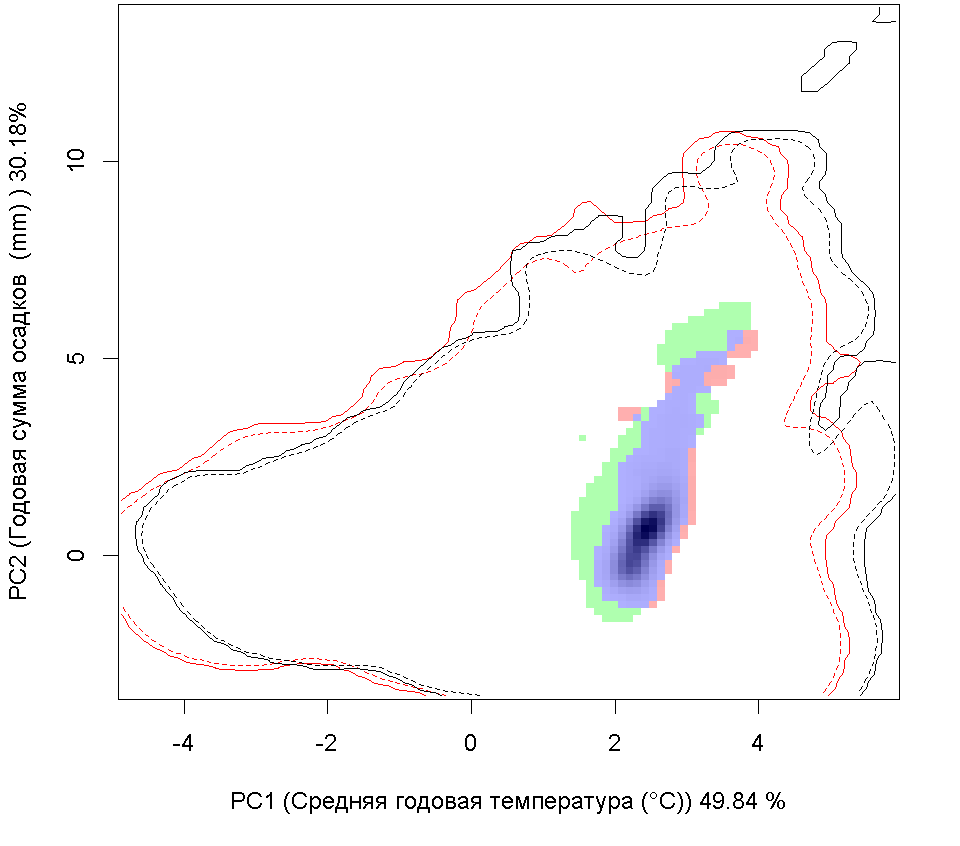 F
F