Представлена версия документа, предназначенная для печати
Вы можете загрузить эту страницу на сайте
Mytilopsis leucophaeata (Conrad, 1831) |
Систематическое положение (Kingdom, Phylum, Class, Order, Family, Species):
Царство — животные, Animalia. Тип — моллюски, Mollusca. Класс — двустворчатые, Bivalvia. Отряд — венероидные, Veneroida. Семейство — дрейссениды, Dreissenidae. Вид — ложная мидия, Mytilopsis leucophaeata
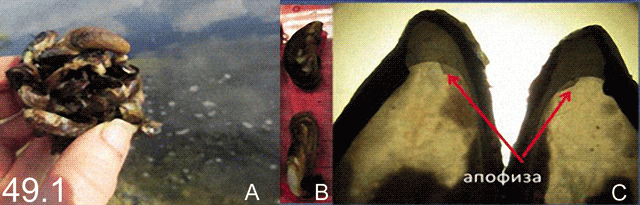
Русское и английское названия. Ложная мидия Cоnrad’s false mussel
Синонимы. Митилопсис, Brackish water mussel, Dark false mussel, Congeria cohleata (Nyst, 1835), Mytilus leucophaeatus Conrad, 1831, Mytilus americanus Recluz, 1858, Mytilus cochleatus Nyst, 1835.
Нативный ареал. Опресненные прибрежные воды Мексиканского залива (Marelli, Gray, 1983)
Современный ареал (мировой и конкретнее в России). Начиная с 1930-х гг. в Новом Свете расселился по Атлантическому побережью Северной Америки, главным образом в эстуариях, нижних и средних течениях крупных рек, на север от нативного ареала дошел до р. Гудзон, на юг до юго-востока Бразилии; найден в нескольких районах юга Новой Англии; встречается и в крупнейшем эстуарии США - Чесапиксиком заливе (Kennedy, 2011; Rizzo et al., 2014).
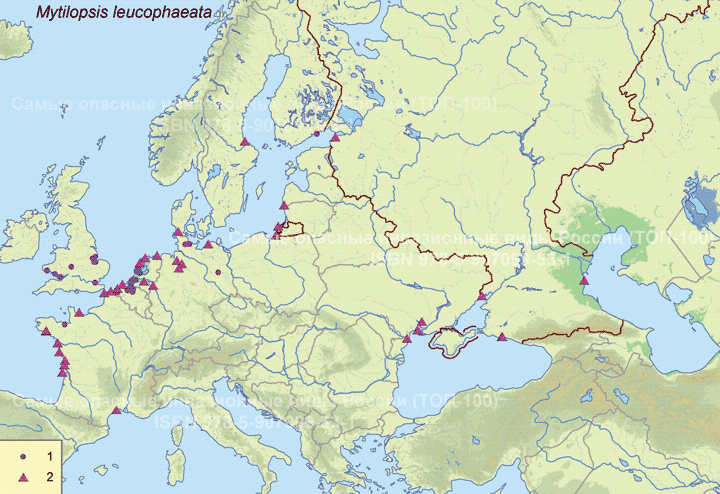
В восточном полушарии обнаружен на западе Африки (Marelli, Gray, 1983). В Европе впервые зарегистрирован в Бельгии (Antwerp Harbor) в 1835 г., расселился в Нидерландах, Франции, Великобритании, Испании, Германии (Forsström et al., 2016). С 1996 по 2015 г. найден в Понто-Каспийском регионе: 5 находок в Черном море и Таганрогском заливе Азовского моря, 3 - в Каспийском море (Zhulidov et al., 2018). В начале XXI в. отмечен в Балтийском море (Финский, Ботнический и Гданьский заливы - прибрежные воды Финляндии, Швеции, России и Польши) (Орлова и др., 2015; Fosström et al., 2016).
В российской части приобретенного ареала зарегистрирован на 3 эстуарных и прибрежных участках Понто-Каспийского региона: в дельте Волги вблизи Астраханского биосферного заповедника (Прикаспий), в Таганрогском заливе (Азовское море) и в бухте р. Туапсе (Черное море) (Zhulidov et al., 2018). В российском секторе Балтийского моря найден в 2014 г., но, судя по размерно-возрастному составу особей, натурализовался здесь до середины 2000-х гг. Популяция пока остается строго в границах прибрежной зоны Копорской Губы Финского залива, отепленной из-за работы Ленинградской атомной станции (Орлова и др., 2015).
Пути(коридоры) и векторы (способы) интродукции. Трансконтинентальное расселение в Европу и Африку могло происходить в балластных камерах судов, тогда как вдоль побережий материков, в эстуариях и нижних течения рек, этот морской эвригалинный прибрежный вид вполне мог расселяться в составе обрастания корпусов судов, барж (Smith, Boss 1996). Есть мнение, что в р. Гудзон мог попасть с обрастанием раковин устриц (Kennedy, 2011).
Как и у прочих беспозвоночных с планктонной личинкой, важную роль в освоении новых экосистем играет саморасселение с течениями, в том числе и с техногенными циркуляционными, вместе с которыми личинки разносятся по системам водоснабжения и охлаждения промышленных объектов и электростанций.
В Черное, Азовское и Каспийское моря интродукция, скорее всего, произошла благодаря судоходству, возможно из портов Западной Европы по европейскому внутриконтинентальному коридору, основу которого составляют реки Дунай и Рейн (Therriault et al., 2004). В Каспий - по Волго-Донскому каналу. В Балтийском море расселение осуществлялось с судоходством, как в балластных водах, так и в обрастании (Darr, Zettler, 2000; Laine et al., 2006).
Местообитание. Ложная мидия обитает в широком диапазоне соленостных условий – от пресных вод до вод соленостью в 25‰ (Verween et al., 2010). Несмотря на то, что в нативном ареале она населяет прибрежные морские воды, в приобретенном зарегистрирована в эстуариях, нижних и даже средних течениях крупных рек (например, Гудзон и Миссиссиппи). В Европе она формирует совместные поселения с понто-каспийским представителем своего семейства - речной дрейссеной (Dreissena polymorpha) на распресненных участках эстуариев (Darr, Zetler, 2000; Therriault et al., 2004). В российской части ареала встречается в олигогалинных и мезогалинных условиях – в Копорской Губе (Балтийском море) при солености 1,2 – 6‰ (Орлова и др., 2015), в Понто-Каспийском регионе при 0,9-10,2‰ (Zhulidov et al., 2018).
Температурные пределы, в которых может обитать - 6,8 -37 ºС (Verween et al., 2010). До недавнего времени считалось, что из-за субтропического происхождения вида, его расселение на севере ареала будет ограничено отепленными участками, что справедливо для популяций Финского залива (Laine, et al., 2006; Орлова и др., 2015), обитающих в тепловом шлейфе сбросных вод атомных станций (в г. Ловиизе, в Финляндии и в Копорской Губе, в России). Однако, находки 2010-х гг. в Финляндии, Швеции и Польше свидетельствуют об адаптации вида и к относительно холодноводным условиям северной и восточной частей Балтийского моря (Forsström et al., 2016).
Ложная мидия, как и дрейссены – обрастатель, прикрепляющийся к поверхностям с помощью биссусных нитей. В природных местообитаниях заселяет как естественные субстраты (камни, раковины других моллюсков и.т.д.), так и искусственные (садки для марикультуры, днища судов, понтоны). Представляет, как и другие дрейссениды, проблему для технических объектов, особенно, если условия благоприятны для теплолюбивых обрастателей. Так в Копорской Губе Финского залива в зоне подогрева и циркуляции водных масс, связанных с работой атомной станции, биомасса поселений ложной мидии достигает 22,7 кг/м2.
Некоторые особенности биологии. Образ жизни такой же, как и у других представителей семейства, демонстрирующих инвазии (дрейссен речной и бугской). Сходен и жизненный цикл. Ложные мидии раздельнополы, самцы и самки выметывают половые продукты в воду, где происходит оплодотворение, а затем из зигот развиваются планктонные личинки (последовательно: трохофора, велигер, поствелигер). Планктонное развитие занимает от 6 до 12 дней в зависимости от температуры воды (Siddall, 1980). Период размножения: от круглогодичного до обычного (с конца весны до начала осени) в умеренных широтах. Моллюски приступают к размножению при прогреве воды до 13-15 ºС (Verween et al., 2010).
Будучи фильтратором, ложная мидия потребляет сестон и можно полагать, что ее роль в экосистемах-реципиентах сходна с таковой обеих инвазионных дрейссен.
Данные о скорости роста противоречивы – от 3-6 мм/год до 15 мм за теплый сезон (Verween et al., 2010), что, скорее всего, как и скорость личиночного развития, связано с температурными и пищевыми условиями конкретного водоема и года.
Влияние вида (на другие виды, экосистемы включая лесную и агроценозы, здоровье человека). Роль ложной мидии в экосистемах изучена недостаточно. Однако можно полагать, что при массовом развитии она близка к описанному для речной и бугской дрейссен.
В Финском заливе с 2017 г. ложная мидия стала пищевым ресурсом бентосоядной рыбы-вселенца - понто-каспийского круглоротого бычка Neogobius melanostomus, популяция которого достигает особенно высокой численности на участке Копорской Губы, плотно заселенном митилопсисом. Очевидно, что планктонные личинки митилопсиса служат пищей другому вселенцу – кордилофоре каспийской, повсеместно распространённой в Копорской Губе.
С хозяйственной точки зрения вид – источник биопомех, особенно для промышленных предприятий и в секторе энергетики, где обрастание может приводить к отказам оборудования (Rajagopal et al., 2002).
Контроль численности. Соблюдение норм обращения с балластными водами рассматривается как превентивная мера против дальнейших интродукций ложной мидии (GISD, 2018). Раннее выявление и своевременное удаление очагов расселения может предотвратить натурализацию в водоемах и облегчить защиту оборудования технических объектов от обрастания (Verween et al., 2010). В искусственных местообитаниях механическое удаление митилопсиса с поверхностей признано неэффективным (Davidson et al., 2008). Рекомендовано применение химических средств для обработки водных потоков, с которыми личинки митилопсиса разносятся по системам, и тщательный подбор минимальной эффективно концентрации и режима обработок, обеспечивающего с одной стороны экономное расходование реагента и минимальное воздействие остаточных концентраций на окружающую среду, с другой - гарантирующее отсутствие репарации повреждения у обработанных индивидуумов в период чистой воды. Оптимален «пульсирующий режим», при котором даже кратковременная обработка воды гипохлоритом натрия с концентрацией активного хлора в 0,6 мг/л, направленная против личинок, дает вполне удовлетворительные результаты (Rajagopal et al., 2002).
Авторы.Орлова М.И., Фенева И.Ю.
|
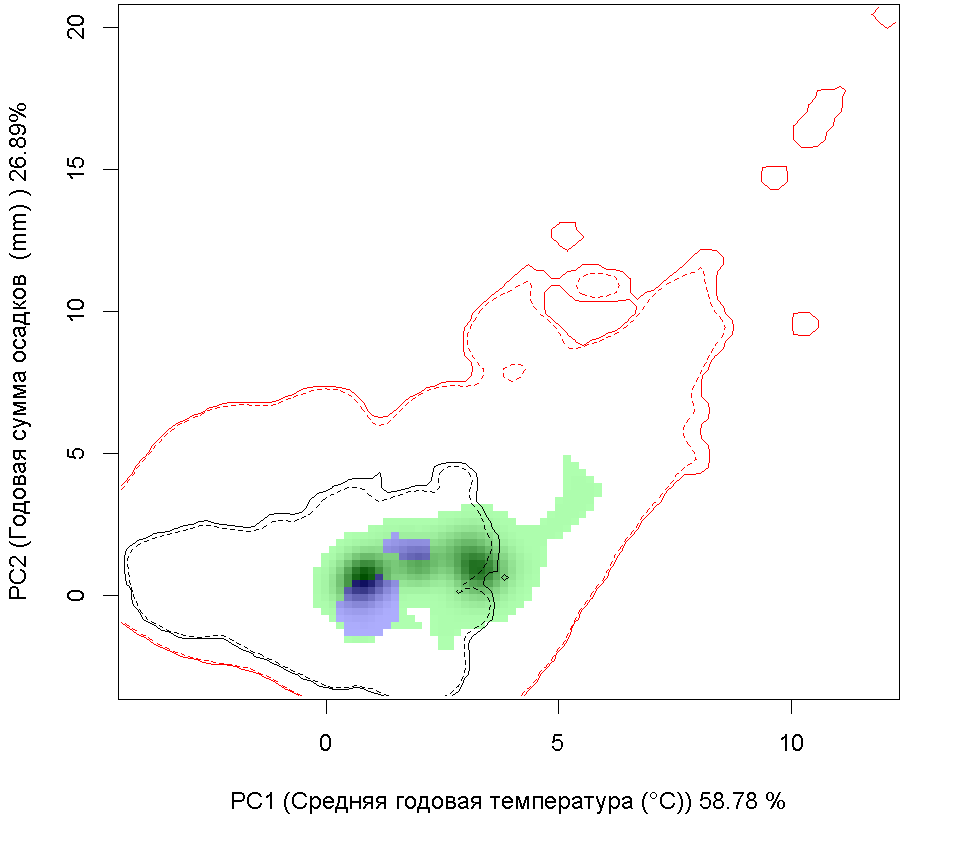
Графическое представление перекрывания ниш нативной и инвазионной частей ареалов, а также при сценариях изменения климата |
|
|
Текущий климат |
|
|
Нативная часть |
Инвазионная часть |
|
|
|
| Графическое представление перекрывания ниш нативной (A) и инвазионной (B) частей ареалов вида, где сиреневый цвет – зона стабильности, розовый цвет – зона расширения, зеленый цвет – зона «неиспользования». Сплошные и пунктирные линии показывают 100% и 90% области доступной среды в нативной (черные линии) и инвазионной (красные линии) частях ареалов, которые использовались для анализа перекрывания ниш. | |
|
Сценарии изменения климата |
|
|
RCP26 |
RCP45 |
|
|
|
|
RCP60 |
RCP85 |
|
|
|
| Графическое представление перекрывания ниш в условиях текущего климата и при сценариях его изменения - (C) RCP26; (D) RCP45; (E) RCP60; (F) RCP85. | |
Литература.
| |
кабинетом «Биоинформатики и моделирования биологических процессов» ИПЭЭ РАН в рамках выполнения гранта РНФ № 16-14-10323. | |
| © 2004-. Кабинет "Биоинформатики и моделирования биологических процессов" ИПЭЭ РАН | |
 A
A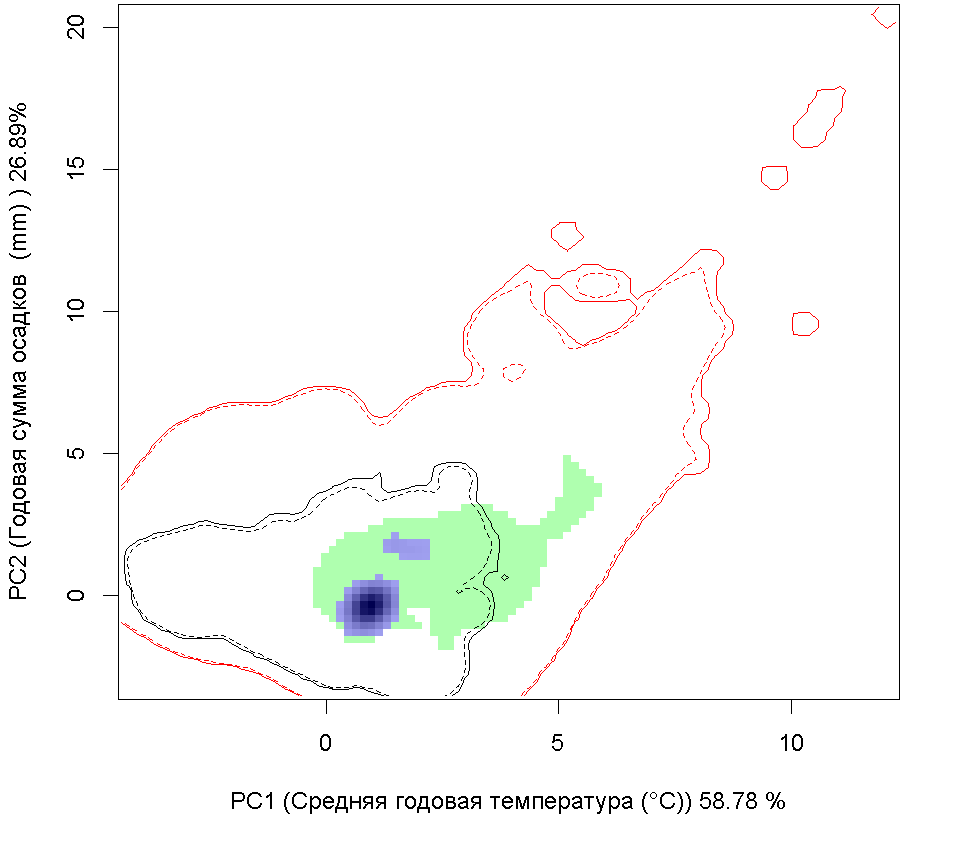 B
B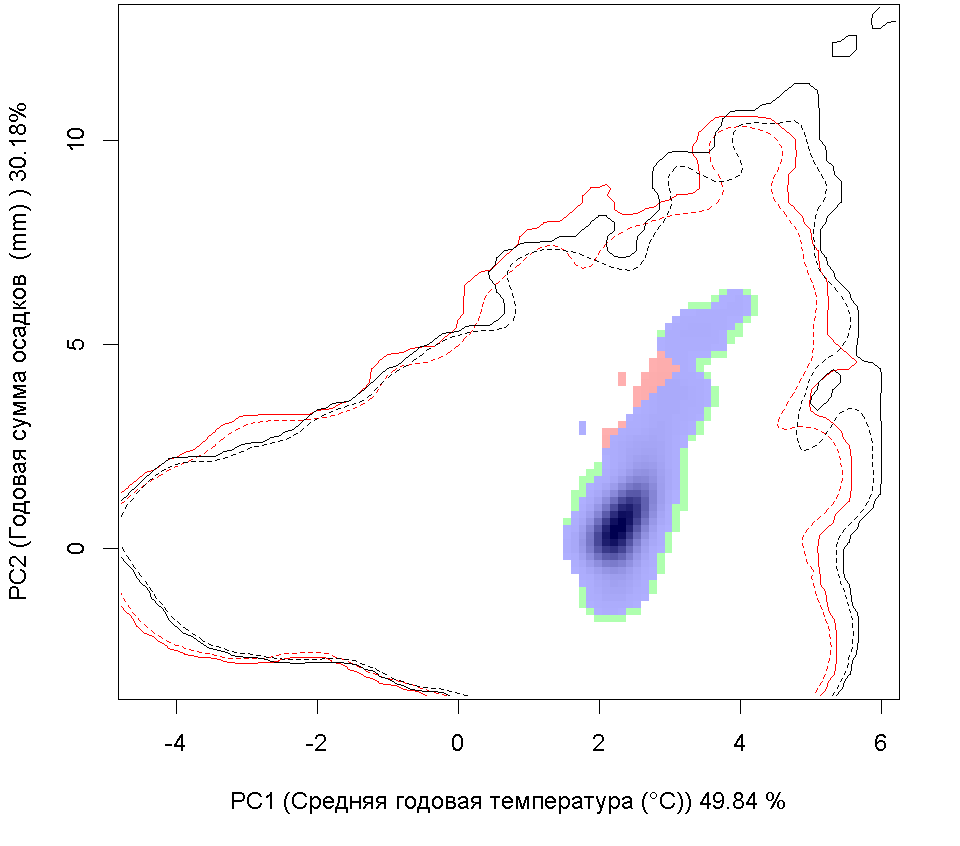 C
C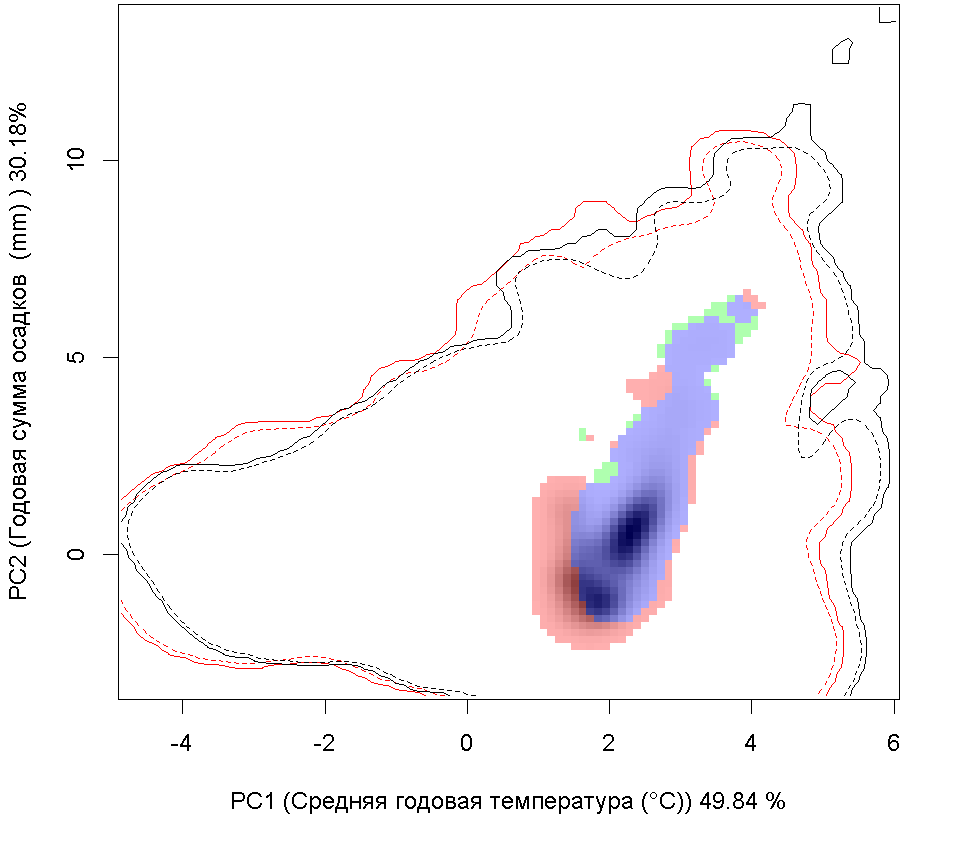 D
D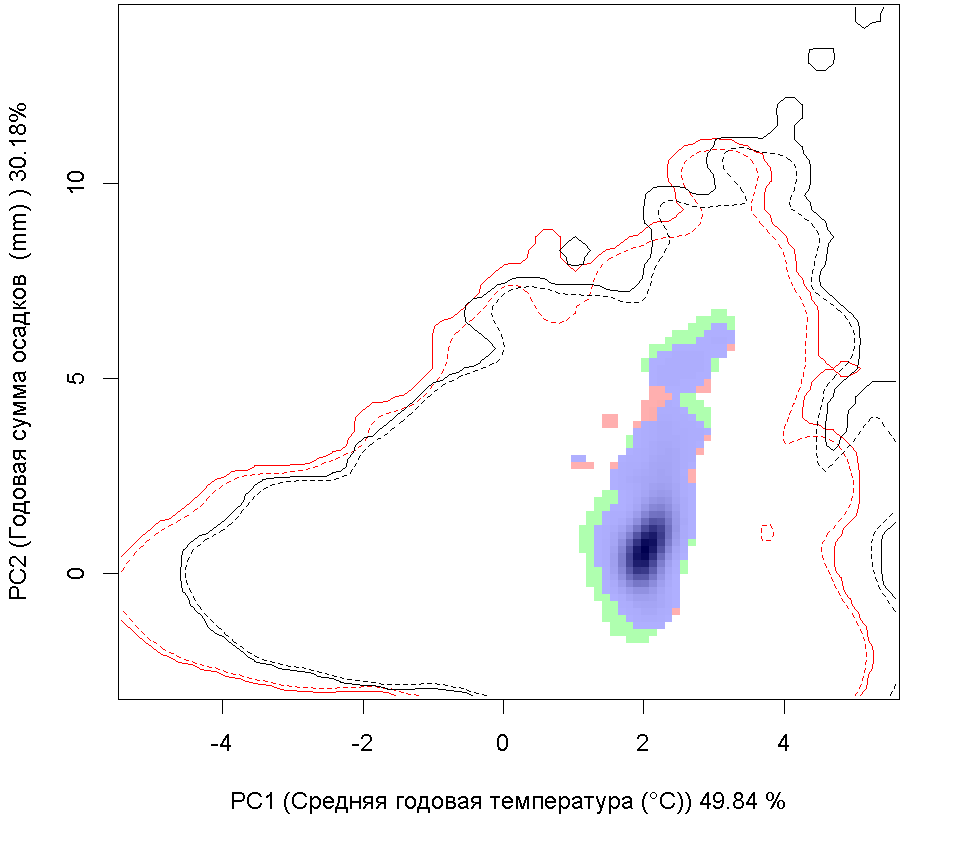 E
E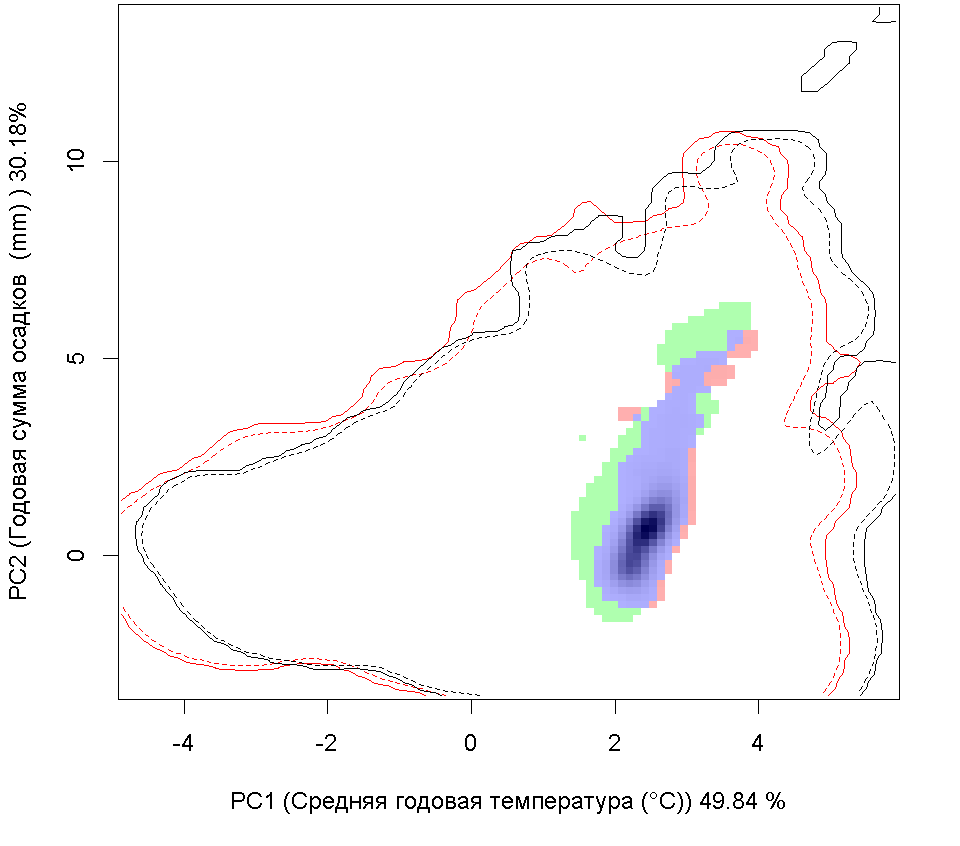 F
F