
|
|
|
|
|
|
Систематическое положение (Kingdom, Phylum, Class, Order, Family, Species): Chromista >> Bacillariophyta >> Bacillariophyceae >> Bacillariales >> Bacillariaceae >> Pseudo-nitzschia calliantha  Псевдонитшия - Pseudo-nitzschia calliantha Lundholm, Moestrup & Hasle Русское и английское названия. Псевдонитшия - Pseudo-nitzschia Синонимы. нет Нативный ареал. Побережье Тихого океана Современный ареал (мировой и конкретнее в России). Морская диатомовая водоросль Pseudo-nitzschia calliantha имеет широкий современный ареал: Арктика: Остров Гершель (Lundholm et al., 2003). Европа: Шотландия: Великобритания (Lundholm, 2003). Черное море (Lundholm et al., 2003, Besiktepe et al., 2008, BSPC, 2014), Хорватия (Lundholm, et al., 2003, Buric et al., 2008), Дания (Lundholm, et al., 2003, Lundholm et al., 2012), Франция (Kipoga, 2006), Греция (Moschandreou et al., 2010), Италия (Lundholm, et al., 2003, Caroppo и др. 2005), Норвегия (Lundholm, et al., 2003), Португалия (Churro et al., 2009), Румыния (Lundholm, et al., 2003), Словения (Lundholm, et al., 2003), Испания (Lundholm, et al., 2003), Турция (Европа) (Bargu et al., 2002). Атлантический океан: Бермудские острова (Lundholm, Моструп, Hasle и Hoef-Эмден, 2003). Северная Америка: Канада, Флорида (Lundholm et al., 2003), Мэриленд (Lundholm et al., 2012.), Нью-Брансуик (Lundholm et al., 2012.), Северная Каролина (Lundholm et al., 2012.), Новая Шотландия ( Lundholm, et al., 2003), Остров принца Эдуарда (Lundholm, et al., 2003). Южная Америка: Бразилия (Eskinazi-Leca et al., 2010), Чили (Lundholm et al., 2003, Alvarez et al., 2009.). Африка: Тунис (Sahraoui et al., 2011, Lundholm et al., 2012..). Азия: Россия: юго-восточные берега России (залив Петра Великого Японского моря, Охотоморское побережье о. Сахалин) (Orlova et al. 2008), Россия (Дальний Восток) (Stonnik et al., 2011.). Юго-Восточной Азии: (Teng et al., 2013) Малайзия, Вьетнам (Lundholm, et al., 2003, Hasle&Lundholm 2005, Lundholm et al., 2006). Австралия и Новая Зеландия: Австралия (McCarthy 2013), Новый Южный Уэльс (Lundholm, et al., 2003, Ajani et al., 2013), Тасмания (Ajani et al., 2013). 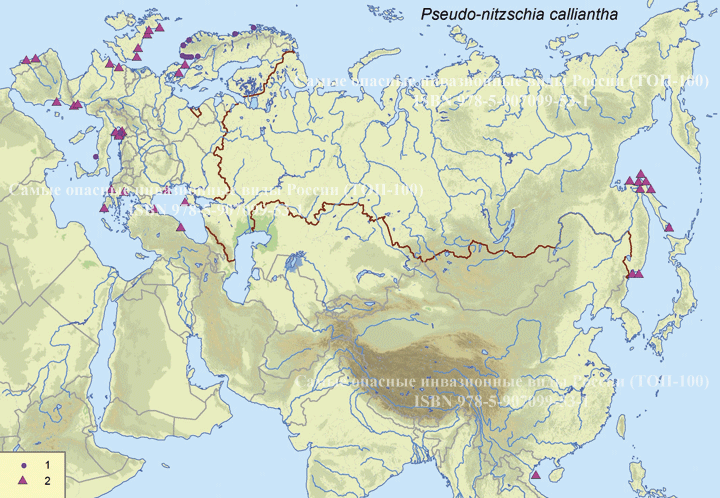 Инвазионная часть ареала псевдонитшии каллианты (Pseudo-nitzschia calliantha) в Евразии. Места находок по 1: (25 October 2018) данным GBIF Occurrence Download https://doi.org/10.15468/dl.xowuhn; 2- литературным данным (см. текст) © Проект РНФ, № 16-14-10323 (Рук. В.Г. Петросян), ИПЭЭ РАН Пути(коридоры) и векторы (способы) интродукции. Распространение с морскими течениями. Местообитание. Морская планктонная водоросль. Побережья морей, лагуны, заливы, а также обнаружена в открытых частях морей и океанов. Особенности биологии. Pseudo-nitzschia calliantha – это нитчатая водоросль, панцирь отдельных клеток имеет удлиненную форму. Кремневые стенки клеток утолщены, что снижает их плавучесть. Это дает им преимущество в том, что, погружаясь в глубину, они избегают губительных уровней освещения, а также выедания зоопланктоном. Клетки продуцирует органические вещества в процессе фотосинтеза. Они имеют центральную вакуоль для запасания питательных веществ с целью последующего использования. Эти водоросли могут длительное время переживать условия недостатка биогенов за счет системы рециркулирования неорганического углерода и азота. Этот вид водоросли может продуцировать две покоящиеся стадии. Одна – это покоящиеся споры, которые могут длительное время находиться в состоянии покоя. Они как правило оседают на дно водоема и активизируются при улучшении условий среды. Другая – это покоящиеся клетки, которые быстрее, чем покоящиеся споры, активизируются при повышении концентрации биогенов. Размножение происходит в основном за счет простого деления. При этом дочерняя клетка получает только одну половину материнского панциря, а вторая половина достраивается, в результате чего происходит снижения размеров клеток популяции. Половое размножение происходит, когда образуются пассивные и активные гаметы, которые сливаются, образуя зиготу, из которой происходит формирование ауксоспоры. Переход к половому размножению инициируется, когда концентрация водорослей превышает определенный порог, т.е. процесс полового размножения возможен только при высокой концентрации клеток. Влияние вида (на другие виды, экосистемы включая лесную и агроценозы, здоровье человека). Диатомовые водоросли рода Pseudo-nitzschia широко распространены и нередко доминируют в прибрежных морских водах всех биогеографических зон Мирового океана. Это единственный род диатомовых водорослей, виды которого продуцируют нейротоксичную домоивую кислоту, которая, передается по пищевым цепям и вызывает отравления людей, а также гибель морских животных. Нейротоксин вызывает амнезическое отравление у человека, морских млекопитающих и птиц (Belonget al., 2012). В связи с увеличением потребления морепродуктов и развитием аквакультуры проблемы, связанные с токсичностью диатомовых водорослей, становятся актуальными в России (Вершинин, Орлова, 2008). Начиная с 1992 г., в прибрежных водах Японского моря, а также у берегов о-ва Сахалин, где находятся хозяйства марикультуры и рекреационные зоны, регулярно отмечают случаи "цветения" воды, вызываемое псевдонитшией.
Литература
Другие ссылки http://www.algaebase.org/search/species/detail/?species_id=73897 http://www.rfbr.ru/rffi/ru/project_search/o_280190
| ||||||||||||||||||||||||||||||||||||||||
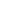

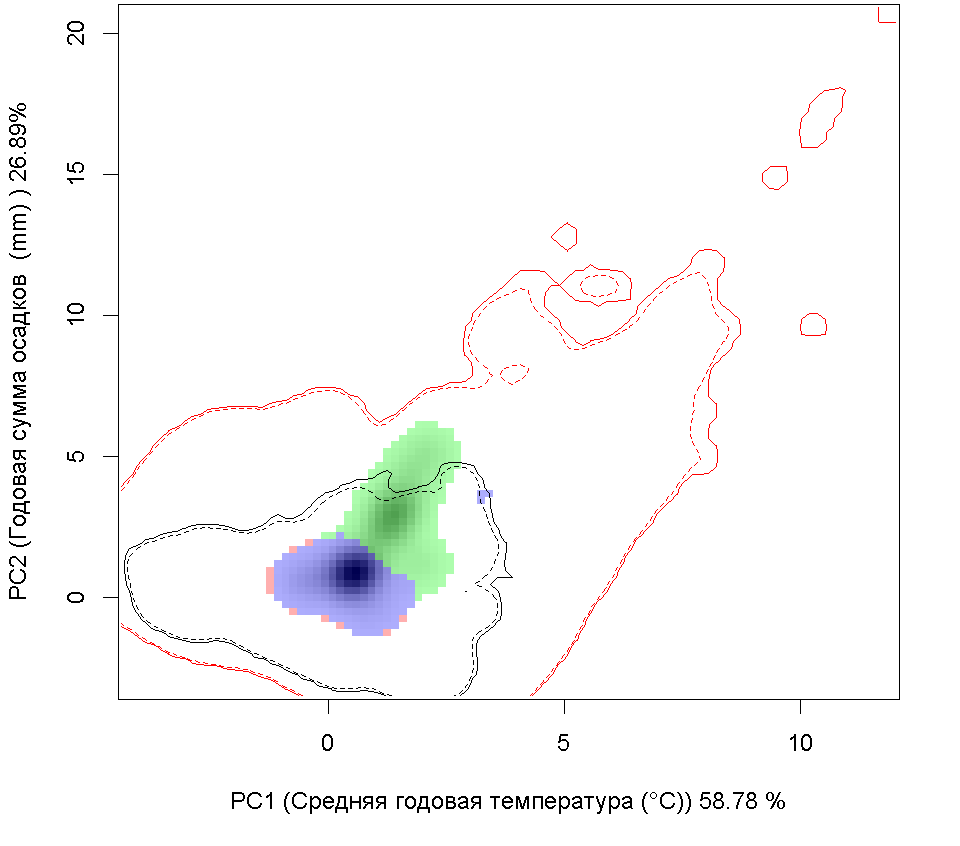 A
A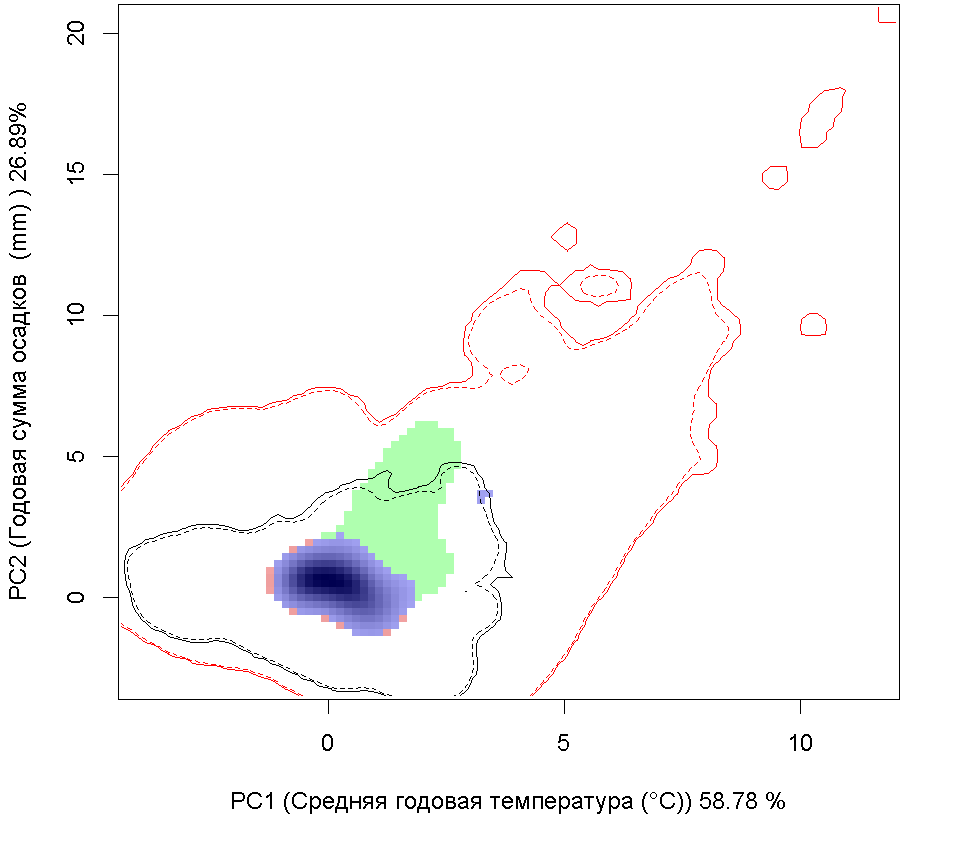 B
B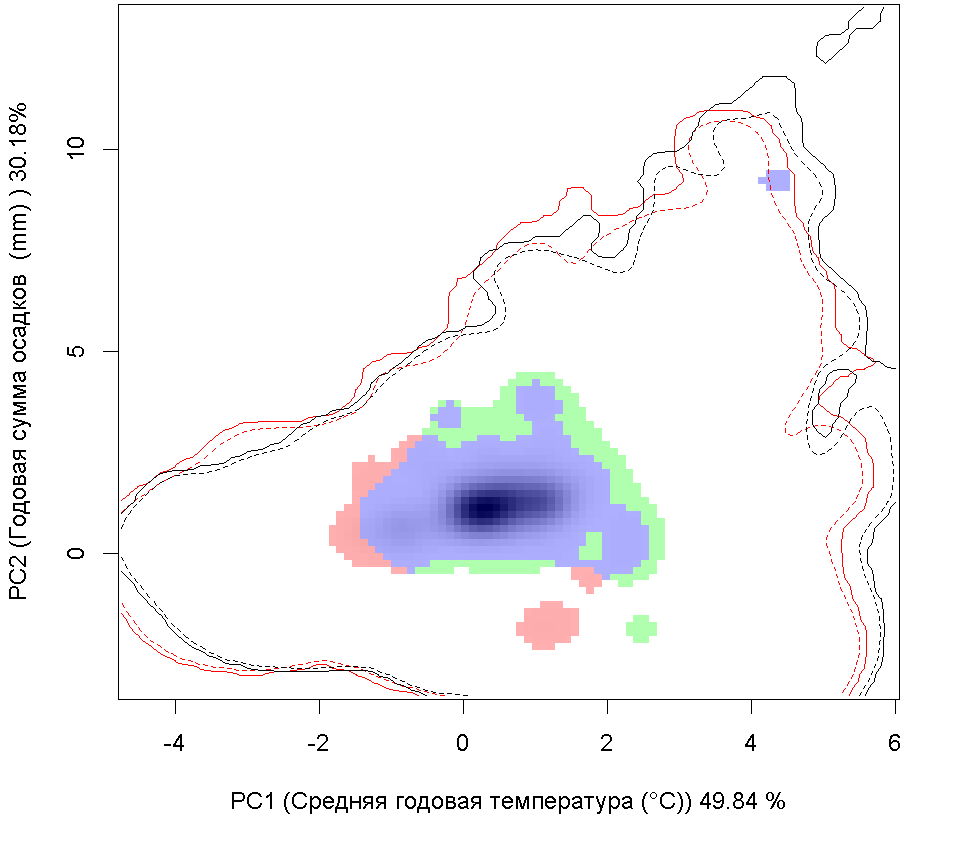 C
C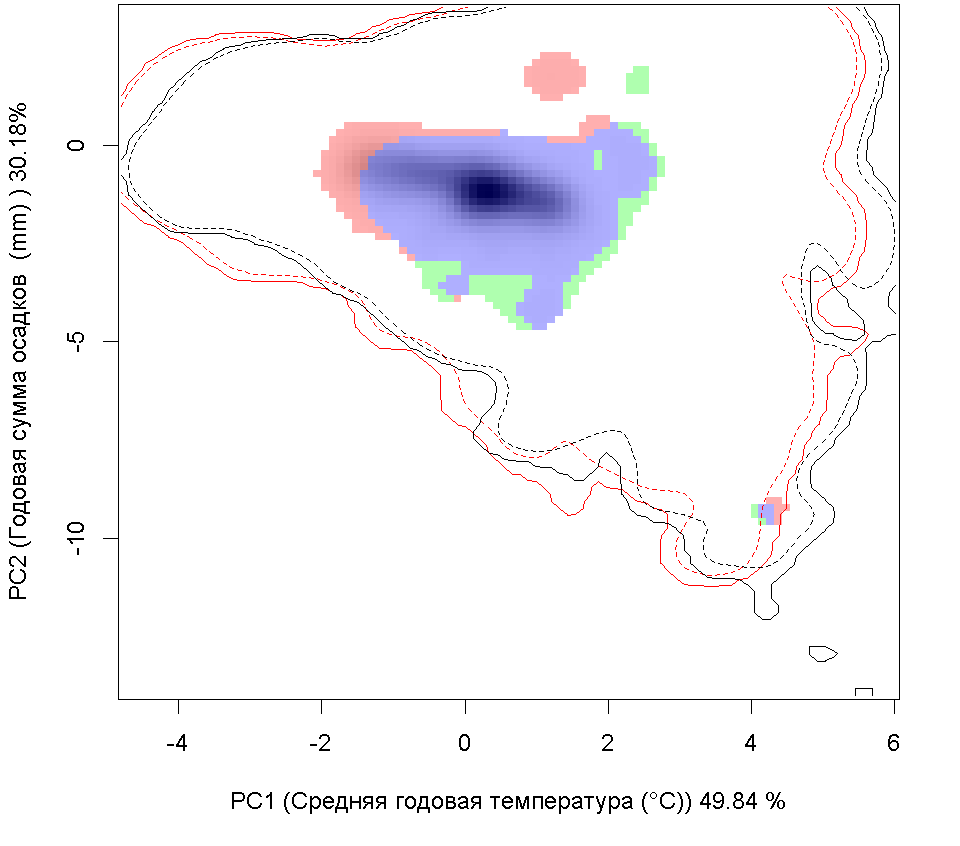 D
D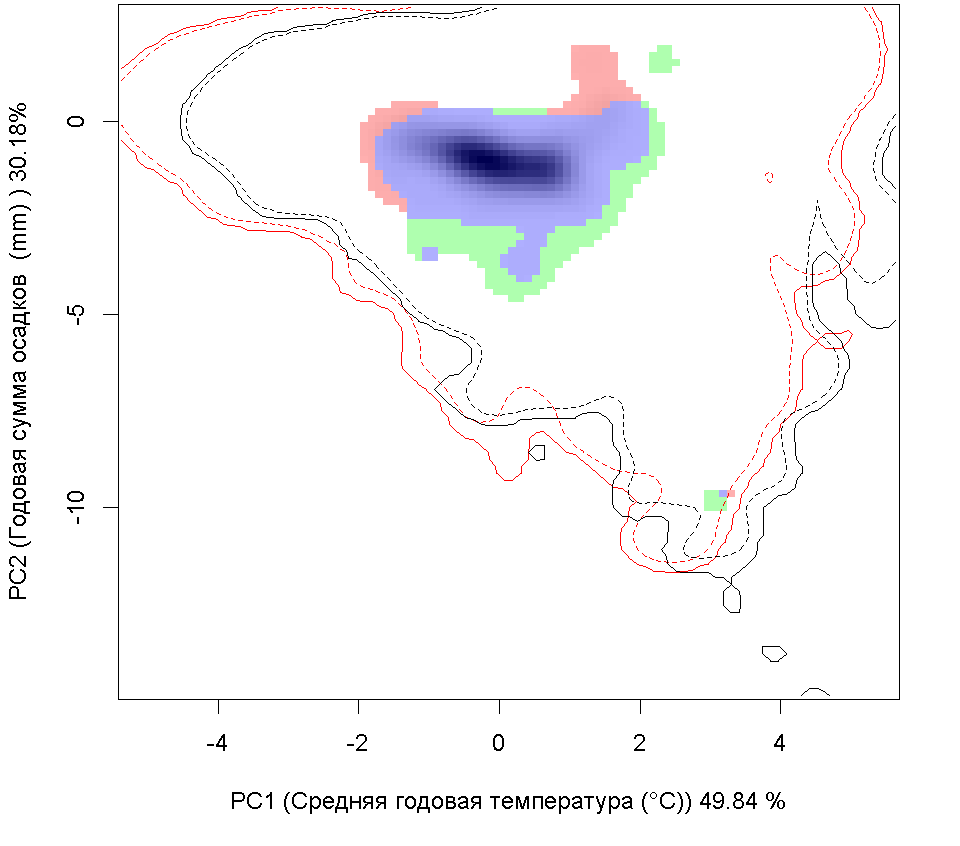 E
E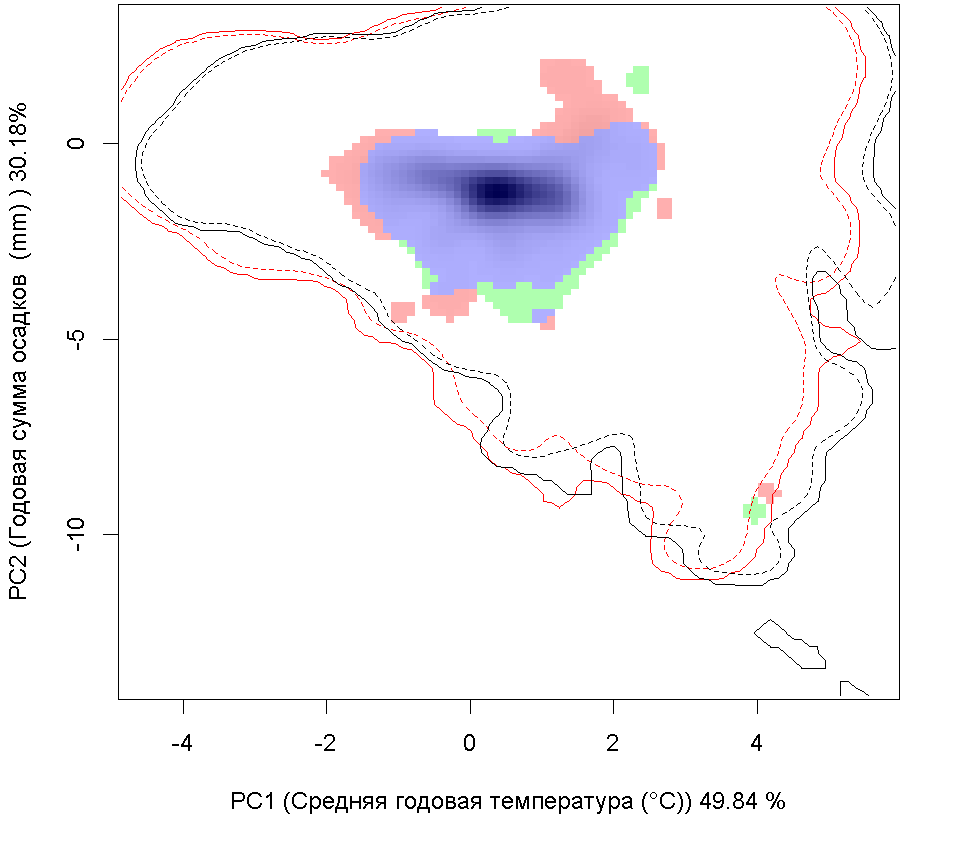 F
F