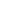|
| |
Oithona davisae Ferrari F.D. & Orsi, 1984
Ойтона |
Систематическое положение (Kingdom, Phylum, Class, Order, Family, Species):
Animalia>> Arthropoda>> Maxillopoda>>Cyclopoida>>Oithonidae>> Oithona davisae
 Ойтона - Oithona davisae Ferrari F.D. & Orsi, 1984
Ойтона - Oithona davisae Ferrari F.D. & Orsi, 1984
Русское и английское названия. Ойтона, Oithona
Синонимы. Нет.
Нативный ареал. Впервые этот вид был описан из Сан-Франциско, штат Калифорния в 1984 году, но считается, что его начальным регионом обитания были воды Японии и Кореи.
Современный ареал (мировой и конкретнее в России).
Этот вид встречается в штатах Калифорния и Вашингтон (США), Испании, Украине и Болгарии, Японии. Встречается у побережья Китая и Вьетнама. В Средиземном море он был обнаружен в 2003 г. Впервые O. davisae была обнаружена в Севастопольской бухте Черного моря в 2005 г. в единичных экземплярах. В следующем году ее численность существенно увеличилась. C 2006 г. Доминирующий вид в зоопланктоне Черного моря в районе Севастополя в летне-осенний период. В 2009 г. ойтона распространилась в вдоль побережья Черного моря, и была отмечена у берегов Болгарии и Румынии. В 2010 г. ойтона была зарегистрирована в северной части Ваттового моря (часть акватории Северного моря).
 Инвазионная часть ареала O. davisae в Европе.
1 – места находок по литературным данным (см. текст)
© Проект РНФ, № 16-14-10323 (Рук. В.Г. Петросян), ИПЭЭ РАН Пути(коридоры) и векторы (способы) интродукции.
Переносится с балластными водами судов.
Особенности биологии. Oithona davisae – раздельнополые животные. Самки крупнее самцов. Они имеют крючковидно изогнутые, укороченные антенны, служащие для схватывания и удержания самок при спаривании. Размер взрослых самок варьирует от 0.54 до 0.62, а самцов от 0.47 до 0.53 мм. К основанию абдомена самок прикреплены два яйцевых мешка. Спариванию предшествует сложное половое поведение, в котором играют важную роль как хеморецепция, так и механорецепция. Самки копепод выделяют половые феромоны, которые воспринимаются самцами при помощи хемосенсорных щетинок первых антенн. При спаривании самец и самка обычно располагаются «головой к хвосту» друг друга. Оплодотворение сперматофорное. Сперматоформ (капсула со сперматозоидами) переносится самцом с помощью задних ножек и приклеивается к генитальному сегменту самок во время спаривания. Из яиц копепод выходит науплиусы, которые проходят пять стадий развития. Шестая стадия называется метанауплиусом, которая является промежуточной между науплиальными стадиями и копеподитами. Науплиусы поедают простейших, бактерий, детрит и компоненты фитопланктона. Особи на стадии науплиусов несегментированные, имеют овальную форму. Они имеют маленький непарный глаз и три пары конечностей. У метанауплиуса имеются зачатки следующей пары конечностей (максилл II). Копеподитных стадий тоже пять. Шестая стадия – взрослые особи. Внешне копеподиты напоминают взрослых особей.
Местообитание.
Обитает в морских, солоноватых и пресных водах. Часто встречается в эсутариях. Относится к теплолюбивым, развиваясь в значительных количествах в тёплый период года. Хорошо приспособлен к жизни в эвтрофных условиях и в прибрежных акваториях. Живет в широком диапазоне солености 3 – 40‰.
Влияние вида (на другие виды, экосистемы включая лесную и агроценозы, здоровье человека).
Всеяднен: питается водорослями, простейшими, коловратками, науплиями. Потенциально может приводить к изменениям в трофических отношениях в экосистемах. Увеличение численности ойтоны и расширение области ее обитания приводит к изменению структуры нативных сообществ и особенностей их функционирования, что было показано на примере модификации зоопланктона и ихтиопланктона Севастопольской бухты и прибрежья Черного моря, вызванной этим вселенцем.
Многочисленные исследования показали, что науплии и копеподитные стадии ойтоны и других мелких копепод служат важным источником пищи личинок рыб и других планктоноядных хищников. С 2006 г. в районе Севастополя отмечена тенденция к улучшению состояния ихтиопланктонного комплекса, что связано с улучшением кормовой базы личинок рыб в результате интродукции ойтоны в Черное море.
|
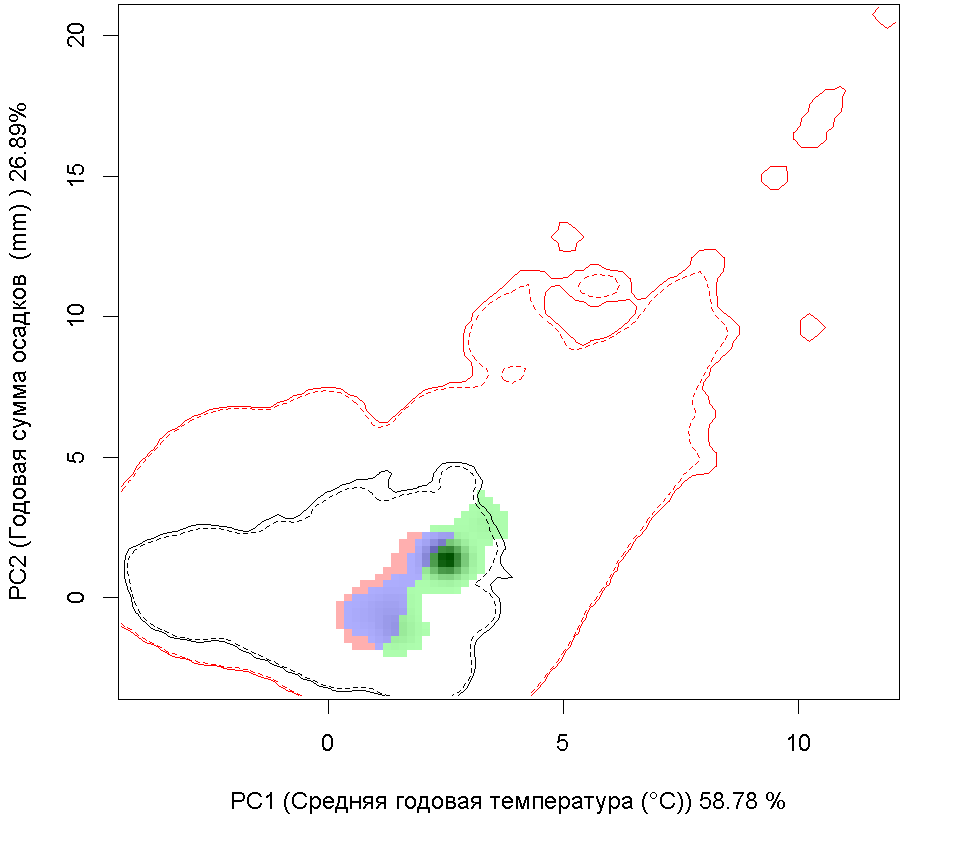
Графическое представление перекрывания ниш нативной и инвазионной частей ареалов, а также при сценариях изменения климата
|
|
Текущий климат
|
|
Нативная часть
|
Инвазионная часть
|
|
 A A
|
 B B
|
| Графическое представление перекрывания ниш нативной (A) и инвазионной (B) частей ареалов вида, где сиреневый цвет – зона стабильности, розовый цвет – зона расширения, зеленый цвет – зона «неиспользования». Сплошные и пунктирные линии показывают 100% и 90% области доступной среды в нативной (черные линии) и инвазионной (красные линии) частях ареалов, которые использовались для анализа перекрывания ниш. |
|
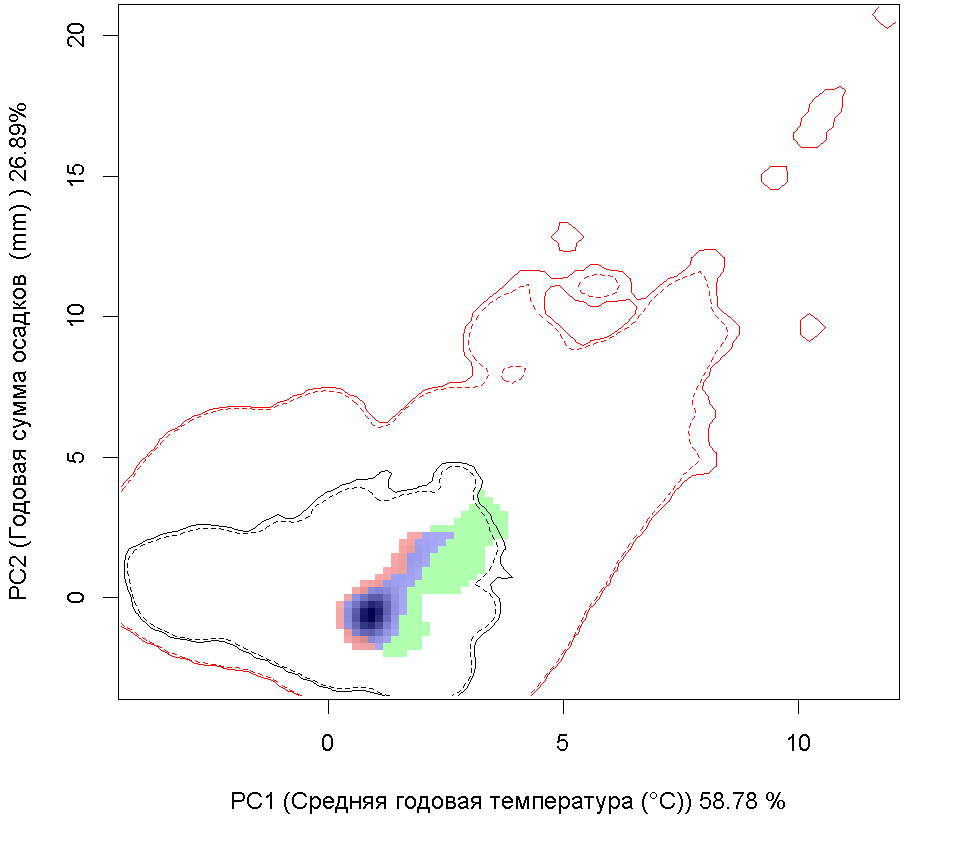
Сценарии изменения климата
|
|
RCP26
|
RCP45
|
|
 C C
|
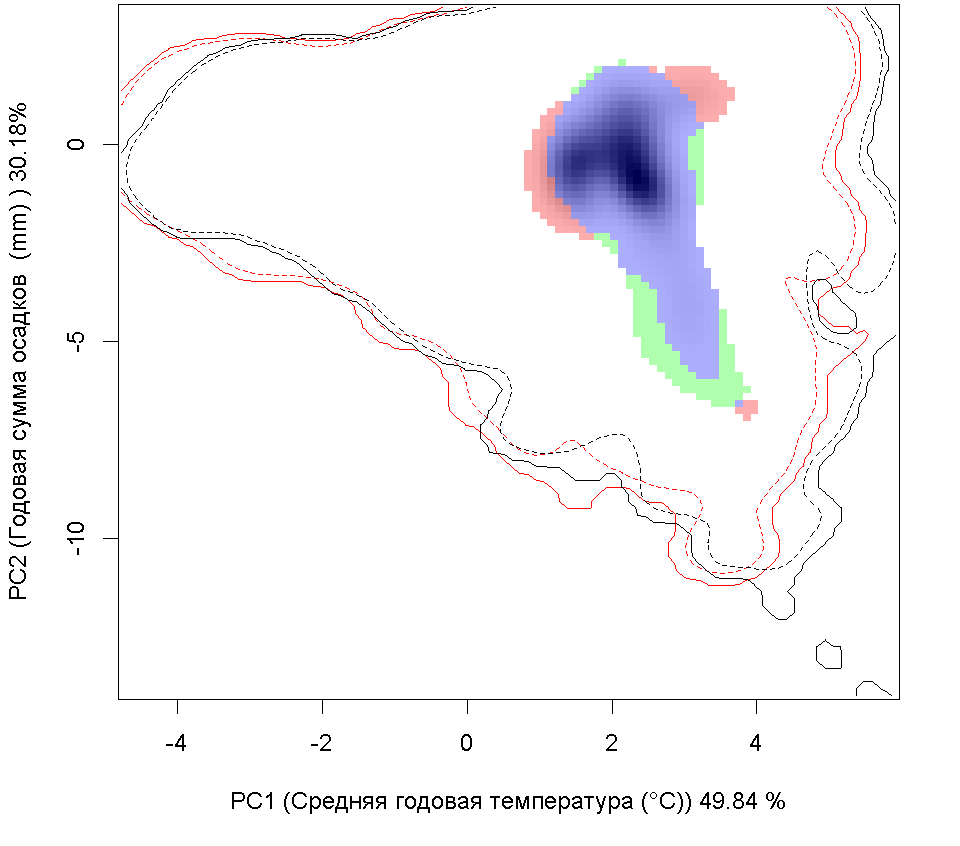 D D
|
|
RCP60
|
RCP85
|
|
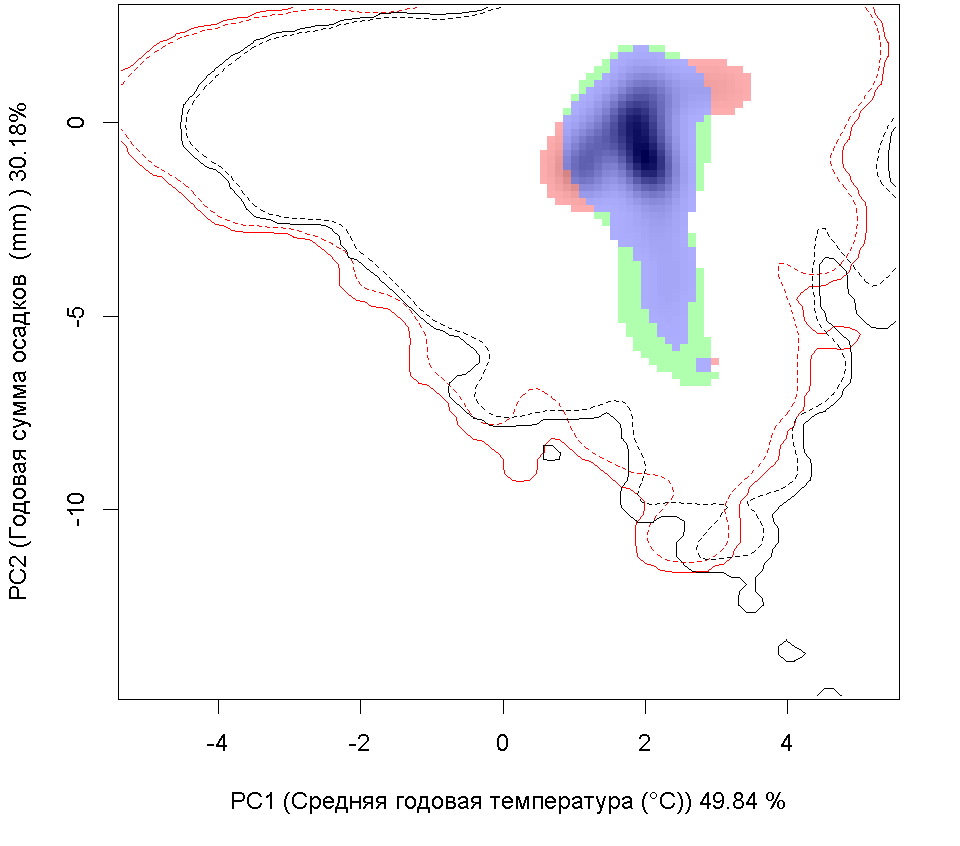 E E
|
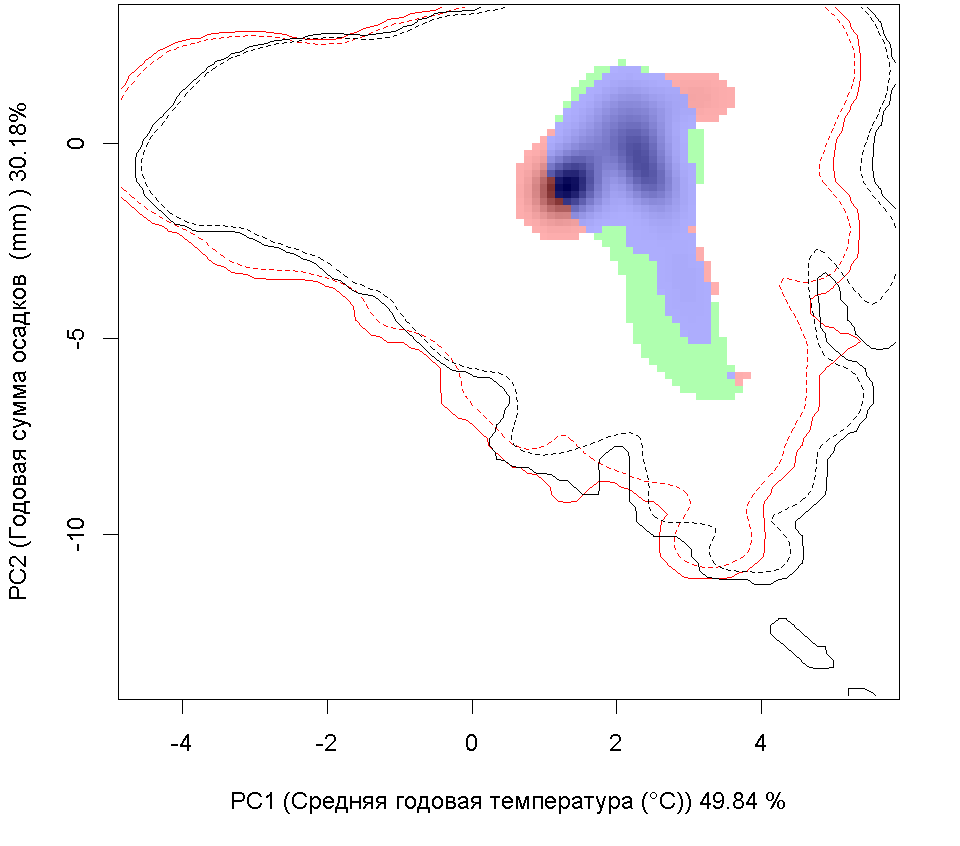 F F
|
| Графическое представление перекрывания ниш в условиях текущего климата и при сценариях его изменения - (C) RCP26; (D) RCP45; (E) RCP60; (F) RCP85. |
Литература
- Серёгин С.А., Попова Е.В. 2016. Многолетняя динамика численности копеподы-вселенца, Oithona Davisae, в прибрежных водах Чёрного моря. Российский Журнал Биологических Инвазий № 3, 2016. Стр. 90-100.
- Almeda, R., Alcaraz, M., Calbet, A. and Saiz, E. 2011. Metabolic rates and carbon budget of early developmental stages of the marine cyclopoid copepod Oithona davisae. Limnology and Oceanography, 56 (1), 403-414. doi: 10.4319/lo.2011.56.1.0403.
- Almeda, R., Alcaraz, M., Calbet, A., Yebra, L. and Saiz E. 2010. Effect of temperature and food concentration on survival, development and growth rates of naupliar stages of Oithona davisae (Copepoda, Cyclopoida). Marine Ecology Progress Series, 410, 97-109. doi:10.3354/meps08625.
- Altukhov, D.A., Gubanova, A.D. and Mukhanov, V.S. 2014. New invasive copepod Oithona davisae Ferrari and Orsi, 1984: Seasonal dynamics in Sevastopol Bay and expansion along the Black Sea coasts. Marine Ecology, 35 (Suppl. 1), 28-34. doi: 10.1111/maec.12168.
- Cornils, A. and Heckmann, B.-W. 2015. First report of the planktonic copepod Oithona davisae in the northern Wadden Sea (North Sea): Evidence for recent invasion? Helgoland Marine Research, 69 (2), 243-248. doi:10.1007/s10152-015-0426-7.
- Gallienne, C.P. and Robins, D.B. 2001. Is Oithona the most important copepod in the world’s oceans? Journal of Plankton Research, 23, 1421-1432. doi:10.1093/plankt/23.12.1421.
- Heuschele, J. and Kiorboe, T. 2012. The smell of virgins. Mating status of females affects male swimming behavior in Oithona davisae. Journal of Plankton Research, 34, 929-935. doi: 10.1093/plankt/fbs054.
- Hirakawa, K. 1988. New records of the North Pacific coastal planktonic copepods, Acartia omorii (Acartiidae) and Oithona davisae (Oithonidae) from southern Chile. Bulletin of Marine Science, 42, 337-339.
- Hiromi, J. 1994. Further studies on respiration of the small planktonic copepod Oithona davisae with special reference to the effect of feeding. Bulletin of the College of Agriculture and Veterinary Medicine Nihon University (Japan), 51, 149-153.
- Hiromi, J., Nagata, T. and Kadota, S. 1988. Respiration of the small planktonic copepod Oithona davisae at different temperatures. Bulletin of the Plankton Society of Japan, 35, 143-148.
- Hubareva, E. and Svetlichny, L. 2013. Salinity and temperature tolerance of alien copepods Acartia tonsa and Oithona davisae in the Black Sea. Rapp. Comm. int. Mer Medit. 40, 742. doi: 10.13140/2.1.1145.3445.
- Isinibilir, M., Svetlichny, L., Hubareva, E., Yilmaz, I.N., Ustun, F., Belmonte, G. and Toklu-Alicli, B. 2011. Adaptability and vulnerability of zooplankton species in the adjacent regions of the Black and Marmara Seas. Journal of Marine Systems, 84, 18-27. doi: 10.1016/j.jmarsys.2010.08.002.
- Isinibilir, M., Kideys, A.E., Tarkan, A. and Yilmaz, N.I. 2008. Annual cycle of zooplankton abundance and species composition in Izmit Bay (the northeastern Marmara Sea). Estuarine, Coastal and Shelf Science, 78, 739-747. doi:10.1016/j.ecss.2008.02.013.
- Lawrence, D. and Cordell, J. 2010. Relative contributions of domestic and foreign sourced ballast water to propagule pressure in Puget Sound, Washington, USA. Biological Conservation, 143, 700-709. doi:10.1016/j.biocon.2009.12.008.
- Mihneva, V. and Stefanova, K. 2013. The non-native copepod Oithona davisae (Ferrari F.D. and Orsi, 1984) in the Western Black Sea: seasonal and annual abundance variability. BioInvasions Records, 2 (2), 119-124. doi: http://dx.doi.org/10.3391/bir.2013.2.2.04.
- Paffenhofer, G.A. 1993. On the ecology of marine cyclopoid copepods (Crustacea, Copepoda). Journal of Plankton Research, 15, 37-55. doi: 10.1093/plankt/15.1.37.
- Saiz, E., Calbet, A., Broglio, E. and Mar?, P. 2003. Effects of small-scale turbulence on copepods: The case of Oithona davisae. Limnology and Oceanography, 48 (3), 1304-1311. doi: 10.4319/lo.2003.48.3.1304.
- Svetlichny, L. and Hubareva, E. 2014. Salinity tolerance of alien copepods Acartia tonsa and Oithona davisae in the Black Sea. Journal of Experimental Marine Biology and Ecology, 461, 201-208. doi: 10.1016/j.jembe.2014.08.012.
- Svetlichny Leonid, Elena Hubareva1, Antonina Khanaychenko1, Alexandra Gubanova1, Denis Altukhov1, Sengul Besiktepe 2016. Adaptive Strategy of Thermophilic Oithona davisae in the Cold Black Sea. Environment. Turkish Journal of Fisheries and Aquatic Sciences 16: 77-90.
- Temnykh, A. and Nishida, Sh. 2012. New record of the planktonic copepod Oithona davisae Ferrari and Orsi in the Black Sea with notes on the identity of “Oithona brevicornis”. Aquatic Invasions, 7, 425-431. doi:http://dx.doi.org/10.3391/ai.2012.7.3.013.
- Uchima, M. 1988. Gut content analysis of neritic copepods Acartia omorii and Oithona davisae by a new method. Marine Ecology Progress Series, 48, 93-97. doi:10.3354/meps048093.
- Uchima, M. and Hirano, R. 1988. Swimming behavior of the marine copepod Oithona davisae: internal control and search for environment. Marine Biology, 99, 47-56. doi: 10.1007/BF00644976.
- Uye, S.-I. and Sano, K. 1995.) Seasonal reproductive biology of the small cyclopoid copepod Oithona davisae in a temperate eutrophic inlet. Marine Ecology Progress Series, 118, 121-128. doi: 10.3354/meps118121.
- Uye, S.-I. and Sano, K. 1998. Seasonal variations in biomass, growth rate and production rate of the small cyclopoid copepod Oithona davisae in a temperate eutrophic inlet. Marine Ecology Progress Series, 163, 37-44. doi: 10.3354/meps163037.
- Zamora-Terol, S. and Saiz, E. 2013. Effects of food concentration on egg production and feeding rates of the cyclopoid copepod Oithona davisae. Limnology and Oceanography, 58 (1), 376-387. doi:10.4319/lo.2013.58.1.0376.
Другие ссылки
http://www.marinespecies.org/aphia.php?p=taxdetails&id=353995
http://invasions.si.edu/nemesis/calnemo/SpeciesSummary.jsp?TSN=88825
http://www.rfbr.ru/rffi/ru/project_search/o_1927424
Карта
Карта
|