
|
|
|
|
|
|
Систематическое положение: Супергруппа (Царство) – альвеоляты, Alveolata. Тип – динофлагелляты, Dinoflagellata. Класс – динофитовые, Dinophyceae. Отряд – пророцентровые, Prorocentrales. Семейство – пророцентрации, Prorocentraceae. Род – пророцентрум, Prorocentrum. Вид – Prorocentrum minimum. 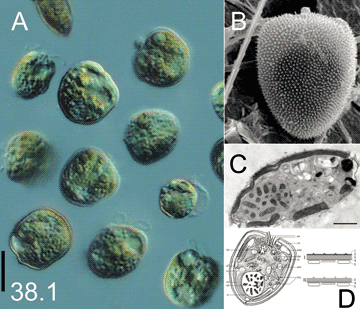 Prorocentrum minimum, жгутиконосцы–динофлагелляты. Живые клетки в культуре (фото М.А. Бердиевой; шкала 10 мкм)(A); клетка в сканирующем электронном микроскопе (фото: M.A. Faust; http://www.vieraslajit.fi/lajit/MX.52909/show) (B); электронограмма (C), (шкала 2 мкм) и схема строения клетки (D) по: Бердиева и др. (2016). Русское и английское названия. Жгутиконосцы–динофлагелляты Dinoflagellates Синонимы. Пророцентрум, Prorocentrum triangulatum Martin, 1929; Exuviaella minima Schiller, 1933; Exuviaella marie-lebouriae Parke & Ballantine, 1957; Prorocentrum cordiformis A.S.Bursa 1959; Prorocentrum marielebouriae (Parke & Ballantine) A.R.Loeblich III, 1970; Prorocentrum cordatum (Ostenfeld) J.D.Dodge, 1975. Нативный ареал. Вид описан из Лионского залива (Средиземное море, южное побережье Франции). Космополит, расширяющий свой ареал. Регион происхождения вида неизвестен. Современный ареал (мировой и конкретнее в России). Распространение всесветное. Особенно широко представлен в северном полушарии в прибрежных водах умеренной и субтропической зоны: в северной части Тихого и Атлантического океанов. В Атлантическом океане – у восточного и южного побережья США: в Чесапикском и Мексиканском заливах; в Карибском море. В северо-восточной Атлантике – от Норвегии до Португалии, включая побережья Северного и Балтийского морей. Зарегистрирован вблизи Великобритании, в проливе Ла-Манш. В Средиземном море известен у берегов Франции, в Адриатическом море. В Черном море впервые найден у берегов Румынии. Обитает в Азовском, Каспийском и Аральском морях (Marazovic et al., 1990). В Тихом океане: у восточных берегов Евразии (от России до Вьетнама, включая Японию, Китай, Тайвань), западных берегов Северной Америки (от Канады до Мексики). Обитает у берегов Филиппинских островов, Австралии и Новой Зеландии. В Дальневосточных морях России зарегистрирован в Беринговом море у берегов Камчатки и в Охотском море; дает наибольшие вспышки цветения в Амурском заливе Японского моря и у берегов Японии (Orlova et al., 2014). В Арктическом бассейне: в Онежском и Двинском заливах Белого моря (Ильяш и др., 2003). Известен в тропических водах у берегов Пакистана; в Оманском заливе Аравийского моря (Al-Hashmi et al., 2015). Учитывая всесветное распространение Prorocentrum minimum, относительно редкая встречаемость этого вида у берегов Африки и Южной Америки объясняется, скорее всего, отсутствием регулярных исследований и мониторинговых наблюдений в этих регионах (Heil et al., 2005).  Распространение Prorocentrum minimum (жгутиконосцы –динофлагелляты) в Евразии. Места находок: 1 - по GBIF.org (21st October 2018) GBIF Occurrence Download https://doi.org/10.15468/dl.ctsyya); 2 – по литературным данным (см. текст); 3 – в акваториях морей. © Проект РНФ, № 16-14-10323 (Рук. В.Г. Петросян), ИПЭЭ РАН Пути(коридоры) и векторы (способы) интродукции. Находки P. minimum в хронологическом порядке следуют так: вдоль румынского берега Черного моря известен с 1950-х гг.; в Северном море впервые обнаружен у берегов Нидерландов в 1976 г., затем у берегов Норвегии; в Датских проливах с 1979 г.; в Балтийском море с 1981 г. В Средиземном море первые цветения зарегистрированы у берегов Франции в 1970 г., с 1983 г. распространен в Адриатическом море. Одна из относительно недавних инвазий P. minimum в Европе – проникновение в Балтийское море. После массового цветения в проливе Скагеррак в 1979 г. этот вид был впервые зарегистрирован в водах западной части Балтийского моря в 1981 г. (Edler et al., 1982). К 1999 г. P. minimum распространился практически по всей акватории Балтики (за исключением Ботнического залива), достигнув на северо-востоке сильно распресненных районов Финского залива (Hajdu et al., 2000, 2005). Наиболее распространенные векторы интродукции: с балластными водами грузовых судов и при интенсификации развития аквакультуры, особенно в условиях эвтрофирования прибрежных вод морей. Местообитание. Эти динофлагелляты обычно встречаются в морских и солоноватых водах в зонах умеренного климата, в субтропических, реже тропических районах. P. minimum присутствует в планктоне во все сезоны года. В основном формирует «цветения» в прибрежных водах морей, в эстуариях и устьях рек, заливах и фьордах, характеризующихся повышенным содержанием биогенных веществ в воде в результате антропогенного воздействия, но может развиваться в больших количествах и в олиготрофных водах пелагиали открытой части морей. Вид с широкой экологической пластичностью: в природе обитает при температуре воды в диапазоне 3-31 оС и солености 2-37‰ (Berland, Grzebyk, 1991; Telesh et al., 2016). Вспышки массового размножения P. minimum («красные приливы») коррелируют с поступлением в воду органических соединений азота (Hajdu et al., 2005). Некоторые особенности биологии.Динофлагелляты Prorocentrum minimum – одноклеточные свободноживущие, планктонные, потенциально токсичные армированные жгутиконосцы. Для этого вида характерна ярко выраженная морфологическая изменчивость (Olenina et al., 2016). Клетки мелкие (длина 14-22 мкм, ширина 10-15 мкм), уплощенные, имеют 2 жгутика, 2 крупные текальные пластинки, которые в виде створок окружают клетку, и 8 мелких апикальных пластинок в зоне жгутикового кармана (Бердиева и др., 2016). Форма P. minimum варьирует от почти круглой или овальной до сердцевидной и треугольной. Вид обладает многими типичными морфологическими, ультраструктурными и биохимическими особенностями, свойственными динофлагеллятам (Околодков, 2011), а также рядом уникальных свойств. Популяции Prorocentrum minimum характеризуются высокими скоростями роста. Клетки обычно размножаются простым делением, однако обнаружена возможность перехода к половому процессу в жизненном цикле P. minimum в условиях дефицита питательных веществ (Skarlato et al., 2018b). Эти жгутиконосцы – преимущественно фотосинтезирующие организмы, но они способны к миксотрофии (Stoecker et al., 1997; Burkholder et al., 2008), т.е. могут питаться как неорганическими соединениями (азотом в виде нитратов, растворенных в воде), так и органическими субстратами (например, мочевиной, которая входит в состав удобрений и попадает в водоем со сточными водами). Если в воде появляется мочевина, то значительная часть клеток P. minimum быстро переключается на ее утилизацию. Обнаружена высокая гетерогенность популяции P. minimum по предпочтению клетками разных питательных субстратов и скорости их потребления; экспериментально установлено ингибирование поглощения нитратов клетками в присутствии мочевины; оценен вклад органических и неорганических субстратов в миксотрофное питание этих инвазийных динофлагеллят, обеспечивающее им преимущество над аборигенными видами в условиях антропогенного эвтрофирования водоемов (Matantseva et al., 2016). Методами физико-химической биологии и водной экологии выявлены адаптационные стратегии P. minimum: продемонстрирована устойчивость жизненного цикла жгутиконосцев к внешним воздействиям и обнаружено повышение синтеза ДНК и РНК клетками в ответ на стресс – изменение температуры и солености воды (Skarlato et al., 2018а). Эти уникальные особенности позволяют вселенцу P. minimum получать конкурентное преимущество над другими видами, в том числе и близкородственными, что способствует его распространению в новые местообитания и вспышкам размножения, приводящим к формированию «красных приливов» (Скарлато, Телеш, 2017). Влияние вида (на другие виды, экосистемы включая лесную и агроценозы, здоровье человека). P. minimum – потенциально токсичные динофлагелляты; они выделяют в окружающую среду продукты жизнедеятельности (вторичные метаболиты), которые могут быть токсичными и опасными для водной биоты и человека. Поэтому вселенец P. minimum, формируя обширные зоны «цветения» в прибрежных водах морей, ухудшает состояние этих экосистем. Высокие концентрации P. minimum в воде и токсичные вещества, накопившиеся в воде в результате цветения этих динофлагеллят, губительно действуют на флору и фауну, аквакультуру, рыболовство, качество вод и здоровье человека. Этот вид вызывает гибель рыб и моллюсков, а также опасен для человека, который потребляет мидии или рыб, отравленных этими жгутиконосцами, или заглатывает воду при купании в водоеме с их массовым развитием (Denardou-Queneherve et al., 1999; Tango et al., 2005). Кроме того, ускорение обмена веществ и энергии в клетках этих простейших как эффективная адаптивная реакция на стресс позволяет им успешно заселять новые прибрежные акватории и процветать в нестабильных условиях, где соленость и качество воды могут меняться сильно и непредсказуемо. Например, заселив Балтийское море, этот вид благодаря своим конкурентным преимуществам вытеснил из состава доминантов близкородственный вид Prorocentrum balticum, который преобладал в микропланктоне до него (Telesh et al., 2016). Всесветное распространение P. minimum позволяет сделать вывод о его высоком инвазионном потенциале и прогнозировать усиление негативного воздействия этого вселенца, снижающее рекреационную и социально-экономическую ценность прибрежных морских экосистем (Heil et al., 2005; Olenina et al., 2010). Контроль численности. Несмотря на то, что вспышки массового размножения динофлагеллят P. minimum зафиксированы в прибрежных водах практически по всему миру, в настоящее время прогнозирование сроков и интенсивности этих «красных приливов» остается проблематичным. Механические, химические и биологические методы борьбы и предотвращения неконтролируемого распространения этого вселенца практически не изучены. Для разработки системы контроля инвазий P. minimum, конкретных мер по ограничению его нежелательного воздействия, прогноза и (в перспективе) регулирования процесса распространения необходимы дальнейшие разносторонние исследования, включающие: - постоянный мониторинг состава и количественного развития фитопланктона, концентрации биогенных веществ и физико-химических параметров в морских прибрежных водах; - контроль распространения этого вида в другие потенциальные водоемы-реципиенты; – изучение адаптивных стратегий вида; - выявление и моделирование условий, при которых P. minimum будет размножаться особенно быстро; – построение валидных карт динамики расширяющегося ареала этого потенциально токсичного вида в морях, омывающих Россию. Авторы Скарлато С. О.,Телеш И. В.
Литература
| ||||||||||||||||||||||||||||||||||||||||

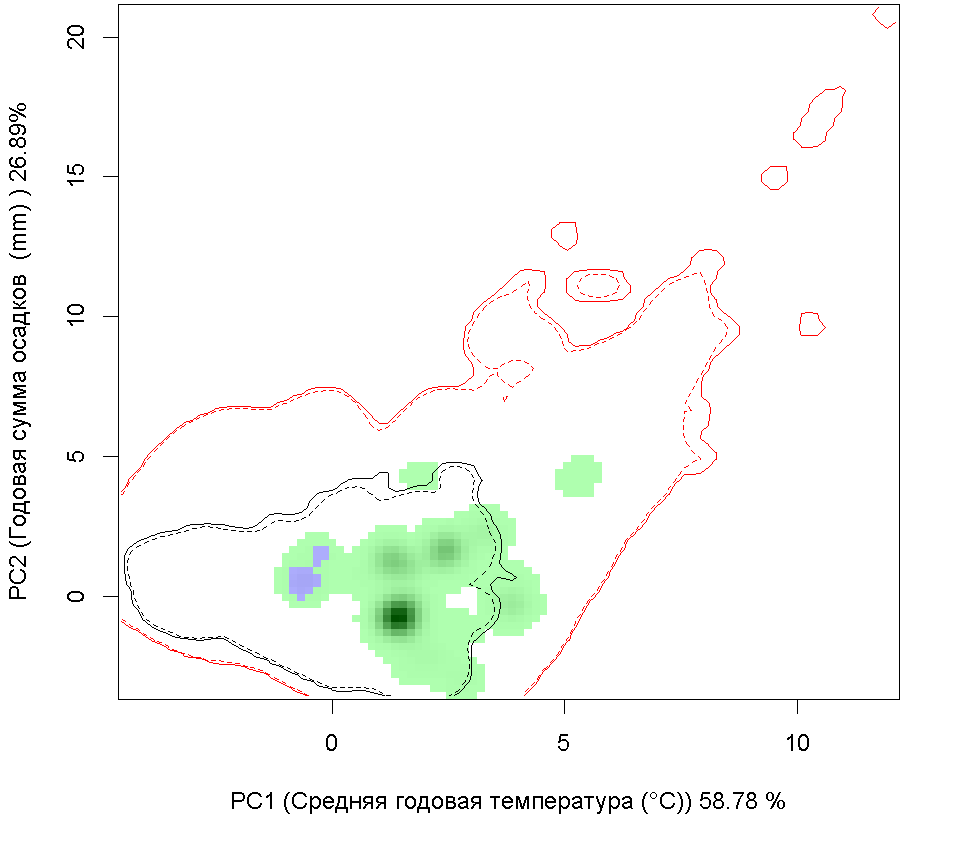 A
A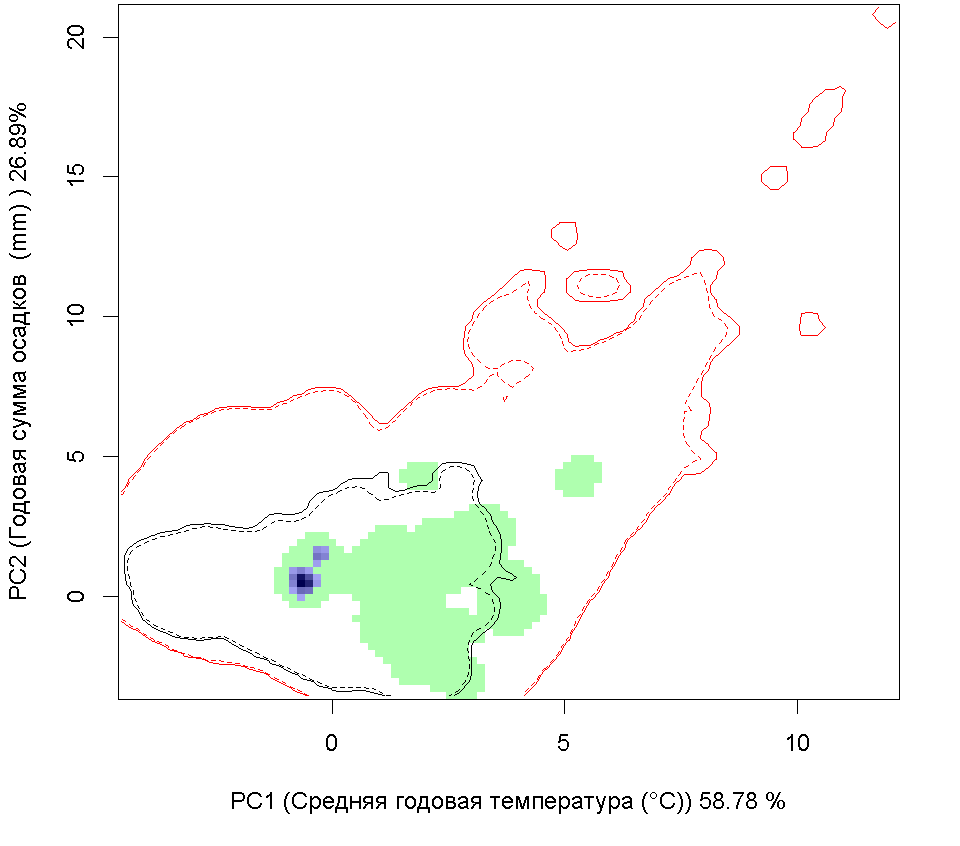 B
B C
C D
D E
E F
F