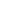|
|
|
|
|
|
Динамика экологических ниш
Для проверки гипотезы консерватизма экологических ниш, которые анализируются здесь в пространстве климатических переменных, и их сравнения в нативной и инвазионной частях ареалов в рамках двух концепций COUE и ECOUE были созданы 100 моделей AENM (для каждого из ИВ, входящего список ТОП-100) в условиях текущего климата. Концепция COUEИтоговые оценки трех мер изменения ниш (стабильности -S, расширения -E, неиспользования-U) и индекс сходства D Schoener’s для видов ТОП-100 приведены в Таблице 1. Индекс стабильности (S) - это доля использования ниш в инвазионной части ареала вида, которая в многомерном экологическом пространстве пересекается с заселенными видом областями в нативной части ареала. Индекс расширения (E) - это доля ниш, освоенных в инвазионной части ареала, неиспользованная видом в нативной части ареала. Индекс не заполнения (U) - это доля ниш, незанятых видом в инвазионной части ареала вида, но которые освоены в нативной части ареала. Поскольку эти индексы оцениваются в пространстве важнейших климатических переменных, определяющих характер распределения видов, то индекс расширения характеризует сдвиги ниш в инвазионной части ареала. Из этой таблицы следует, что по сходству экологических ниш виды в рамках концепции COUE можно разделить на две группы. Для видов первой группы подтверждается гипотеза консерватизма ниш, а для видов второй группы отмечен сдвиг ниш. К первой группе относятся 28 видов, для которых ниши в рамках концепции COUE не различаются(P-value <0.05). Для остальных 71 видов их ниши в нативной и инвазионной частях ареала значимо различаются в концепции COUE. Из этого следует, что вывод о невыполнения консерватизма ниш для 71 вида требует дополнительной проверки в рамках расширенной концепции ECOUE. Концепция ECOUEВ рамках схемы COUE величины E и U оцениваются относительно плотности встречаемости вида в инвазионной и нативной частях ареалах соответственно. Несмотря на то, что S представляет собой перекрытие между нативными и инвазионными нишами, однако его величина оценивается относительно плотности встречаемости вида в инвазионной части ареала и непосредственно рассчитывается как дополнение расширению ниш, т. е. S = 1 - E. Чтобы преобразовать значения индекса в тот же масштаб, в концепции ECOUE величину U оценивают относительно плотности встречаемости в инвазионном ареале с помощью формулы UT=(S/(1-U))/U. Поскольку индексы E, S, UT определены в одном масштабе, то новые модифицирование значения Em, Sm, Um определяются как Em=E/(E+S+UT), Sm=S/(E+S+UT) и Um=UT/(E+S+UT). Далее в концепции ECOUE метрики Em, Sm, Um используются для определения ширины ниш в нативной и инвазионной частях ареала с помощью формул Bn= Sm+Um и Bi= Sm+ Em. Для удобства интерпретации результатов в концепции ECOUE и сравнительного анализа ниш используется ln отношения ширины ниш (breadth ratio, BR), т.е. BR=ln(Bn/Bi). Из этой формулы видно, что если ширины ниш в нативной и инвазионной частях ареала равны Bn=Bi, то BR=0. Если Bn>Bi, тогда BR принимает положительные значения, в противном случае - отрицательные значения. В ECOUE относительное положение ниши в двухмерном пространстве климатических показателей окружающей среды выражается как сходство нативной и инвазионной ниш, а не как расстояние между центроидами ниш. Сходство ниш (Sim) количественно оценивается с помощью Sørensen’s similarity index (Baselga, 2017), т.е. Sim=2Sm/(Bn+Bi). Метрика Sim варьируется от 0 до 1, которые отражают крайние случаи, когда две ниши не перекрываются или перекрываются полностью в пространстве климатических показателей окружающей среды, соответственно. Если две ниши занимают более сходные позиции (т. е. похожие ниши), тогда Sim> 0.5, в противном случае две ниши занимают разные позиции, тогда Sim<0.5. Важно заметить, что сходство ниш в концепции ECOUE отличается от теста на сходство ниш в рамках концепции COUE. В концепции ECOUE принимается важное условие по использованию пороговых значений BR и Sim для индикации сохранения или сдвига ниш. Ниши считаются законсервированными, если либо ниша расширялась медленно, т. е. нативная ниша была больше, чем инвазионная ниша (Pearman et al., 2008), либо положение ниши менялось медленно, т. е. нативная и инвазионная ниши занимали аналогичные позиции в пространстве климатических параметров окружающей среды (Broennimann et al., 2007). Эти пороговые значения в дальнейшем мы использовали для проверки гипотезы консерватизма климатических ниш для пяти экологических групп организмов, на которые были разделены ИВ ТОП-100: «микроорганизмы» (бактерии, хромисты, грибы, нематоды, альвеоляты), растения, наземные эндотермы (птицы и млекопитающие), наземные эктотермы (земноводные, рептилии, насекомые) и водные виды (асцидии, гребневики, моллюски, ракообразные, лучепёрые рыбы). Принимая во внимание основные пороговые значения для BR и Sim в рамках концепции ЕCOUE, можно утверждать, что гипотеза консерватизма справедлива почти для всех исследованных ИВ, и не выполняется только для одного вида, т.е. BR<0 и Sim<0.5 (см. рисунок 29) - Cameraria ohridella (каштановая минирующая моль), для которого ширина инвазионной ниши в 2.3 раза больше нативной, а индекс расширения Em (0.66) существенно больше Sm (0.12) и Um (0.21). Важный вывод из этого анализа заключается в том, что если в рамках концепции COUE гипотеза консерватизма климатических ниш выполнялся для 28 из 99 видов, то в рамках модифицированной концепции ЕCOUE указанная гипотеза выполняется большинства видов за исключением одного. Таблица 1. Оценки трех метрик ниш в рамках концепции COUE (стабильность - S, расширение - E, незаполнение - U) и индекса сходства Schoener’s D для видов ТОП-100
Рисунок 29. Взаимосвязь между сходством и ln-отношением ширины ниш для всех видов (A) и для каждой группы организмов (B-F). Сходство ниш оценивается с помощью индекса сходства Сёренсена (Sim), а размеры ниш с помощью ln-трансформации отношения ширины ниш в нативной и инвазивной частях ареала. На каждой панели точки с ln-отношением ширины ниш больше 0 указывают на то, что нативная ниша больше инвазивной, а сходство ниш больше 0,5 указывает на то, что эти две ниши занимают сходное положение в экологическом пространстве.
С использованием метрик ниш (BR, Sim) на основе построенных AENM моделей показано, что сходство ниш при реализации разных моделей и сценариев изменения климата принимает достаточно высокое значение для всех таксонов (первый этап 2021 г) – бактерии (Sim=0.83±0.21); хромисты (Sim=0.92±0.05); грибы (Sim=0.84±0.14) и растения (Sim=0.92±0.05). Эти высокие значения косвенно подтверждают, что условие консерватизма климатических ниш выполняется для всех видов вне зависимости от модели, сценария и временного периода. В действительности, построенные нами графики взаимосвязи между индексами сходства и ln-отношениями ширины ниш в условиях текущего климата и при реализации разных моделей и сценариев изменения климата для всех видов (рисунок 30) за исключением трех случаев (на рисунке указаны красной стрелкой) иллюстрируют, что, гипотеза консерватизма сохранения климатических ниш в пространстве и времени принимается с вероятностью 0.997 (=1-3/1176).
Рисунок 30. Взаимосвязь между сходством и ln-отношением ширины ниш для всех 37 видов при реализации различных моделей и сценариев изменения климата в период 2020-2100 годов с шагом 20 лет. Сходство ниш оценивается с помощью индекса сходства Сёренсена (Sim), а размеры ниш с помощью ln-трансформации отношения ширины ниш в условиях текущего климата и ожидаемой ниши при изменении климата. На каждой панели точки с ln-отношением ширины ниш больше 0 указывают на то, что текущая ниша больше ожидаемой ниши при изменения климата, а сходство ниш больше 0,5 указывает на то, что эти две ниши занимают сходное положение в экологическом пространстве. Проверка гипотезы климатических ниш в рамках концепции ECOUE. С использованием метрик ниш (BR, Sim) в рамках расширенной концепции с использованием AENM моделей показало, что сходство ниш при реализации разных моделях и сценариях изменения климата принимает достаточно высокое значение для беспозвоночных животных (второй этап проекта, 2022 год) – альвеолят (Sim=0.92±0.06); гребневиков (Sim=0.90±0.24); нематод (Sim=0.94±0.05), моллюсков (Sim=0.68±0.21), ракообразных (Sim=0.68±0.2) и насекомых (Sim=0.69±0.24). Эти высокие значения косвенно подтверждают, что условие консерватизма климатических ниш выполняется для многих видов вне зависимости от модели, сценария и временного периода. В действительности, построенные нами графики взаимосвязи между индексами сходства и ln-отношениями ширины ниш в условиях текущего климата и при реализации разных моделей и сценариев изменения климата для всех видов (Рис. 31) за исключением 79 случаев (на рисунке указаны стрелками) иллюстрируют, что, гипотеза консерватизма сохранения климатических ниш в пространстве и времени принимается с вероятностью 0.95 (=1-79/1488). Важно отметить, что отклонения от гипотезы чаще наблюдаются у насекомых (67 случаев из 79, 84,8 %). Это означает, что в отличие от насекомых представители других таксонов (альвеолят, гребневиков, моллюсков, ракообразных) более консервативны и склонны сохранять экологические ниши при глобальном потеплении.
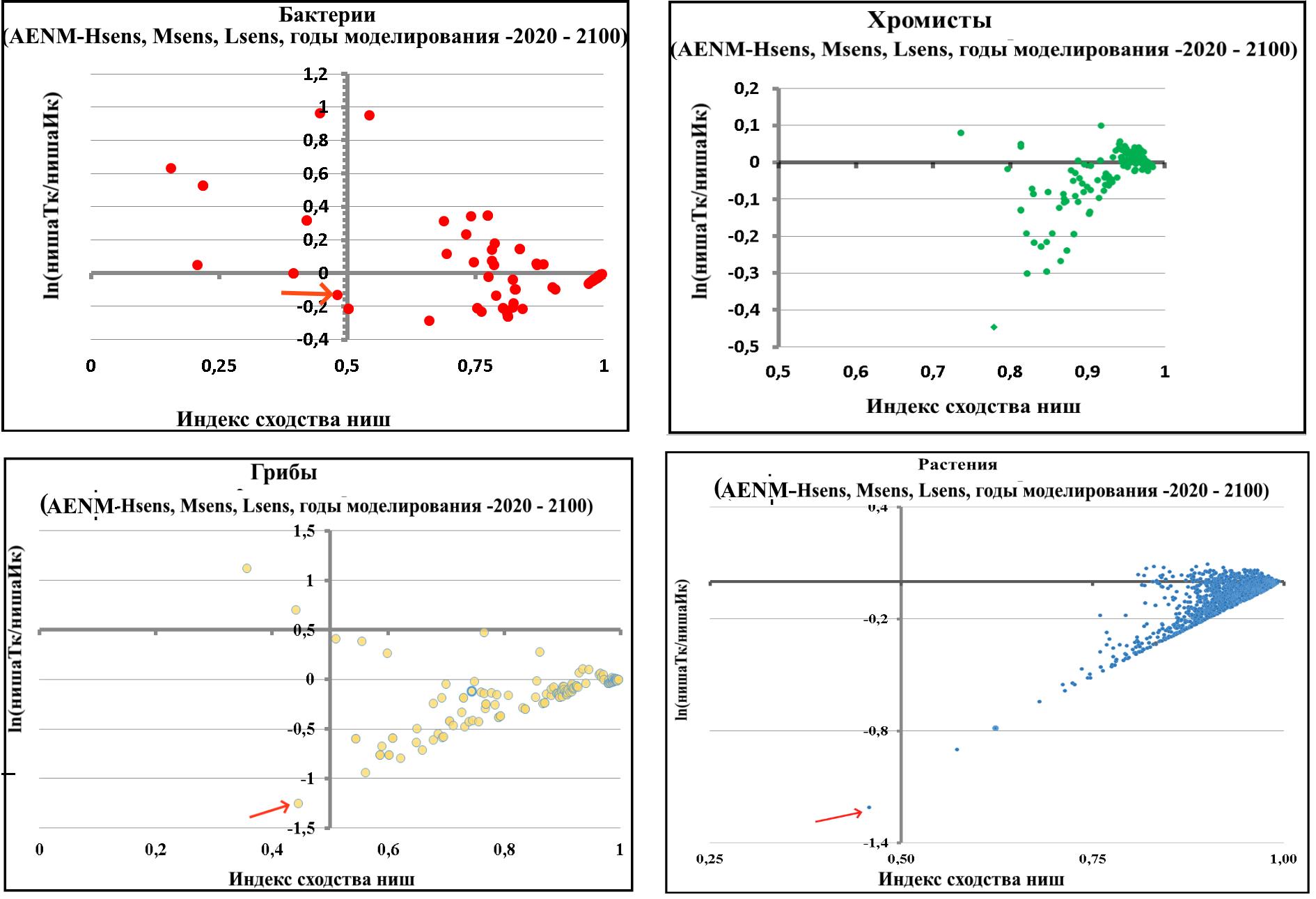
Рисунок 31. Взаимосвязь между сходством и ln-отношением ширины ниш для 43 видов при реализации различных моделей и сценариев изменения климата в период 2020-2100 годов с шагом 20 лет в рамках концепции ECOUE. Сходство ниш оценивается с помощью индекса сходства Сёренсена (Sim), а размеры ниш с помощью ln-трансформации отношения ширины ниш в условиях текущего климата и ожидаемой ниши при изменении климата. На каждой панели точки с ln-отношением ширины ниш больше 0 указывают на то, что текущая ниша больше ожидаемой ниши при изменения климата, а сходство ниш больше 0,5 указывает на то, что эти две ниши занимают сходное положение в экологическом пространстве.  Рисунок 32. Взаимосвязь между сходством и ln-отношением ширины ниш для всех 19 видов при реализации различных моделей и сценариев изменения климата в период 2020-2100 годов с шагом 20 лет в рамках концепции ECOUE. Сходство ниш оценивается с помощью индекса сходства Сёренсена (Sim), а размеры ниш с помощью ln-трансформации отношения ширины ниш в условиях текущего климата и ожидаемой ниши при изменении климата. На каждой панели точки с ln (Bn/Bi)-отношением ширины ниш больше 0 указывают на то, что текущая ниша больше ожидаемой ниши при изменении климата, а сходство ниш S больше 0.5 указывает на то, что эти две ниши занимают сходное положение в экологическом пространстве. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
кабинетом «Биоинформатики и моделирования биологических процессов» ИПЭЭ РАН в рамках выполнения грантов РНФ № 16-14-10323, 21-14-00123. | |
| © 12 май, 2016 - . Кабинет "Биоинформатики и моделирования биологических процессов" ИПЭЭ РАН | |